Using the system for the social preference test in C57BL/6J mice
Figure 1 shows three versions of the experimental set-up. The first version (Figure 1A-C) is designed for mice with dark fur colors, such as C57BL/6J mice. The second (Figure 1D-F) is planned for mice with bright fur colors, such as BALB/c or ICR (CD-1) mice. The third is larger (Figure 1G-I) and designed for rats with bright fur colors, such as SD rats. The first subjects examined were naïve adult (8-12 weeks old), group-housed (two to five animals per cage) male C57BL/6J mice. Social stimuli were juvenile (21-30 days old), group-housed (two to six animals per cage), male C57BL/6J mice. Object stimuli were odorless plastic toys with distinct colors (insets in Figure 1C,I). Video files of the experiments were analyzed using the TrackRodent software (body-based algorithm, see exemplified screen picture and GUI in Figure 2A), which automatically and continuously tracked subject location based on 1) body center and 2) the contact of its body contours with the stimuli-containing chambers.
The analysis involved the following procedures. After uploading a movie file and graphically defining areas to be excluded from the analysis, the experimenter graphically defined two areas, each comprising a distinct chamber, as 'stimuli' (Figure 2A, yellowish areas). Users can also define up to five areas as virtual 'compartments'. Then, users choose the "BlackMouseBodyBased23_7_14_Fast" algorithm from of the list of possible algorithms (Figure 2B). After pressing the Start analysis button, the software tracked the animal location frame-by-frame and depicted the mouse location trace in the arena (Figure 2A, blue line). During this analysis, the software also tracked contacts between the 'stimuli' areas and body contour of the subject (Figure 2C). Such contact events were defined as investigatory and served to calculate the investigation time for each stimulus and the time during which no investigation behavior was displayed (Figure 2D). As apparent in Figure 2C,D showing the analyzed movie, higher investigation time for 'stimulus 2 was observed', which was the social stimulus, compared to the object, which in this case was defined as 'stimulus 1'.
Population analysis of the social preference test in C57BL/6J mice
After allowing the TrackRodent software to analyze all video files of the experimental group, a population analysis for this group was performed using the "TrackRodentPopulationSummary" GUI (Figure 2E), while choosing all possible analyses of this software. The distinct outputs of these analyses for the SP test performed with 58 C57BL/6J adult male mice are displayed in Figure 3, exactly as they appeared on the computer screen, with all error bars depicting SEM. Statistical analyses of these results were previously published11.
The mean total investigation time of the population of subjects towards each of the two stimuli (same-sex juvenile social stimulus and an object) is displayed in Figure 3A, which depicts a clear preference of the subject towards the social stimulus. Although the algorithm is simple, an excellent correlation was observed (r2 = 0.91, p < 10-6, Pearson's correlation) between the investigation time manually measured by a trained observer and the value calculated by the software. The dynamics of this preference can be seen in Figure 3B, in which the mean investigation time for each stimulus is plotted along time in 20 s bins. As shown, social preference is maintained by the subjects throughout the session, although it seemed slight stronger in early stages.
Figure 3C depicts the investigation time for each stimulus, categorized according to bout duration into short (≤6 s), medium (>6 s, ≤19 s), and long (>19 s) bouts, along with the total investigation time. The distribution of investigation time for each stimulus according to bout duration in 1 s bins is displayed in Figure 3D. As shown in Figure 3C,D, there was no difference between stimuli in the short bouts, while the medium and long bouts showed clear social preferences. These results suggest that in C57BL/6J mice, short bouts reflect curiosity, while long bouts reflect an interaction between the subject and stimuli. The relative duration of investigation (RDI, [social-object]/[social+object]) values for each category (short, medium, and long bouts), as well as for the total investigation time, are shown in Figure 3E. As shown, the highest RDI value is achieved with the long bouts, suggesting that they best reflect the social preferences of subjects.
To analyze the dynamics of investigation bout during the test, distributions over time for each category were separately plotted. The distributions over time of the short (Figure 3F) and medium (Figure 3G) investigation bouts suggest a gradual reduction of these bouts over time. In contrast, a similar analysis of the long bouts (Figure 3H) shows a gradual increase over the course of the test. This tendency is also reflected by the distribution of the mean bout duration (Figure 3I), showing a gradual increase over time. Thus, it seems as if the subjects exhibited curiosity mainly during the early stages of the test, while later they displayed more interactions with the stimuli. It should be noted that during the last minute, there is an inherent bias of the analysis towards short bouts, because long bouts were artificially terminated at the end of the session. Therefore, when considering the time course of the investigation bouts, the last minute should be ignored.
We also analyzed the intervals between successive investigation bouts towards the same stimulus (i.e., the time it took the subject to return to the same stimulus after the end of each investigation bout). This parameter reflects the urge of the subject to return to the same stimulus and investigate. Here, the intervals were also categorized into short (≤5 s), medium (>5 s, ≤20 s), and long (>20 s) bouts, which are shown together with total time intervals for each stimulus in Figure 3J. The histogram of the intervals in 1 s bins in displayed in Figure 3K. As shown in Figure 3J,K, the intervals between investigations of the social stimulus were much shorter than those between investigations of the object. Accordingly, the RDI values of intervals in the long range yielded the highest absolute value (Figure 3L); hence, they may be used as an investigative time-independent variable assessing social preference. The distributions over time of intervals plotted separately for each category (Figure 3M-O) suggest a gradual reduction in short and medium intervals and an opposite change in long intervals between investigation bouts, which most likely reflect the general lengthening of investigation bouts. Similar to the distribution of investigation bouts, the last minute of intervals is biased towards short values and should be ignored.
We also analyzed the transitions observed in subjects from one stimulus to another. This plot is displayed in Figure 3P, at which every timepoint the subject started to investigate a new stimulus is marked by a blue dot, every row represents a distinct subject, and the red line represents the mean transition rate. As apparent, the transition rate was highest (~1.5 transitions/min) during the first 2 min of the test and was gradually reduced to less than 50% of the peak. The same tendency is apparent from the distribution of mean transition rate over time (Figure 3Q).
Finally, heatmaps of investigation bouts towards the social stimulus (Figure 3R) and object (Figure 3S) during the course of the test for each subject are shown, with the colors marking bout duration (Figure 3S). Altogether, the results suggest that in C57Bl/6J mice, the SP test is roughly divided into an early exploratory phase (characterized by a high transition rate and short investigation bouts) and a late interaction phase (characterized by low transition rate and long investigation bouts).
Population analysis of the social novelty preference test in SD rats
Figure 4 shows the same analysis as above for the SNP test performed by 59 SD rats. Typically, the preference of novel vs. familiar social stimuli is weaker than the preference towards a novel social stimulus vs. an object, exhibited by mice in the SP test (Figure 3). Nevertheless, the social novelty preference is reflected clearly by all parameters. Notably, as concluded above from the SP test in mice, the long investigation bouts and long intervals are the parameters most clearly exhibiting differences between the stimuli, as reflected by their highest absolute RDI values (Figure 4E,L; it should be noted that RDI values for the SNP test are defined as [familiar-novel]/[familiar+novel]).
Loss of whiskers impairs social preference in both C57BL/6J mice and SD rats
To evaluate the effects of somatosensory manipulation on the dynamics of social behavior, analysis was performed of the SP test completed by 29 C57BL/6J mice and 33 SD rats following whisker trimming (conducted 3-7 days before testing). Then, these results were compared to control animals. As displayed in Figure 5A-F, the whisker-trimmed mice did not prefer the social stimulus over the object, as reflected by the lack of difference in investigation time between the two stimuli (compare Figure 5A auf Figure 5D) and the loss of long investigation bouts towards to social stimulus (compare Figure 5B auf Figure 5E). Interestingly, the whisker-trimmed mice also showed increased rates of transition compared to control mice (compare Figure 5C auf Figure 5F).
Altogether, these data suggest that whisker-trimmed mice markedly reduced the duration of their interactions with the social stimulus. In contrast to mice, whisker-trimmed rats did not lose their social preferences (Figure 5G-L). However, they significantly changed their behavior during the first minute of the test, when they had fewer long investigation bouts (compare Figure 5H auf Figure 5K) and many more transitions (compare Figure 5I auf Figure 5L). Thus, whisker trimming significantly modified the behavior of both rats and mice in the SP test, but in a very different manner between the two species. These results suggest a distinct role for whisker-dependent somatosensory stimulation in social interactions of rats and mice.

Figure 1: Experimental set-up. (A) A schematic depiction of the experimental arena designed for mice with black fur. (B) A picture of the arena from above, using dim red light, showing a C57BL/6J subject mouse in the arena. (C) A picture showing the meshed area of a white chamber, through which the subject interacts with a stimulus (inset: a picture of an object stimulus used for mice). (D-F) As shown in panels A-C, an experimental arena and chambers designed for mice with bright fur. (G-I) As shown in panels A-C, an experimental arena and chambers designed for rats with bright fur (inset in [I]: picture of an object stimulus used for rats). Please click here to view a larger version of this figure.
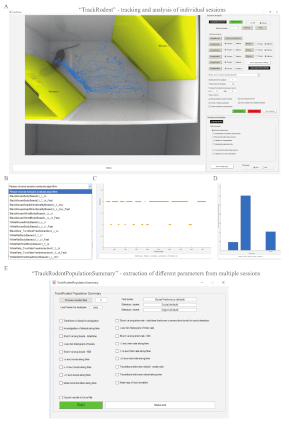
Figure 2: TrackRodent software. (A) A screen of a computer running the TrackRodent software. Left: the video image at the beginning of the analyzed movie (first frame chosen by the experimenter), showing the labeled 'stimuli' areas and tracked movement path of the C57BL/6J subject. Right: the software GUI. (B) The list of optional codes of the software to be used with the GUI. Each of these codes fits a distinct experimental condition. (C) The analysis of the specific experiment shown in panel A, when the option of Stimuli exploration along session was selected in the Results presentation section of the GUI. This plot shows (at each frame) whether the subject was in contact with 'stimulus 1' or 'stimulus 2'. In the presented case, a clear preference towards 'stimulus 2' is evident by the higher number and duration of investigation bouts detected towards this stimulus. (D) The analysis of the experiment shown in panels A and C, when the option of Total stimuli exploration time was selected in the Results presentation section of the GUI. Here too, a clear preference towards 'stimulus 2' is evident from the higher level of investigation time towards this stimulus. (E) The GUI of the TrackRodentPopulationSummary software that enables analysis and plotting of the results from a population of animals, each of which is analyzed using the TrackRodent software (see Figure 3 for the results of such analysis). Please click here to view a larger version of this figure.
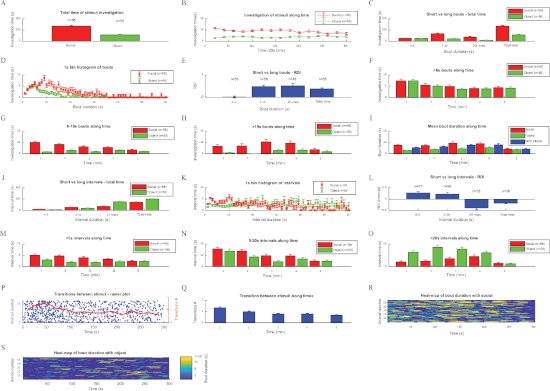
Figure 3: Analyzing the investigation behavior of C57BL/6J mice during SP test using the TrackRodent software. This figure shows the various plots of all optional analyses of the TrackRodentPopulationSummary software (Figure 2E) employed for a population of 58 C57BL/6J adult male mice performing the SP test. The various plots are displayed in a similar order and manner as they appeared on the computer screen (see the representative results section for a detailed explanation of each plot). (A) Total time of stimuli investigation, (B) Investigation of stimuli along time, (C) Short vs long bouts – total time, (D) 1s bin histogram of bouts, (E) Short vs long bouts – RDI, (F) <6 s bouts along time, (G) 6−19 s bouts along time, (H) >19 s bouts along time, (I) Mean bout duration along time, (J) Short vs long intervals – total time, (K) 1 s bin histogram of intervals, (L) Short vs long intervals – RDI, (M) <5 s intervals along time, (N) 5−20 s intervals along time, (O) >20 s intervals along time, (P) Transitions between stimuli – raster plot, (Q) Transition between stimuli along times, (R) Heat-map of bout duration with social, (S) Heat-map of bout duration with object. Please click here to view a larger version of this figure.
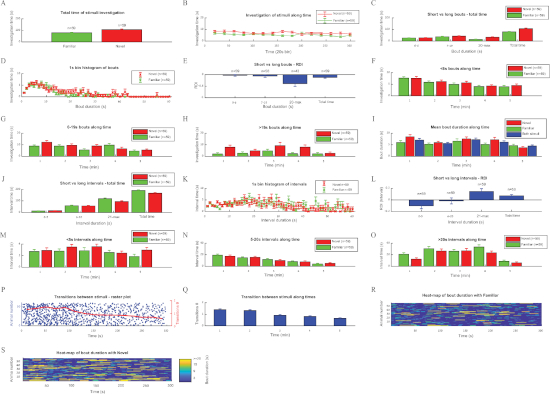
Figure 4: Analyzing the investigation behavior of SD rats during SNP test using the TrackRodent software. Plots for all optional analyses of the TrackRodentPopulationSummary software (Figure 2E) employed for a population of 59 SD adult male rats performing the SNP test. The various plots are displayed in a similar order and manner as they appeared on the computer screen (see the representative results section for a detailed explanation of each plot). (A-Q) Same as described in Figure 3. (R) Heat-map of bout duration with Familiar, (S) Heat-map of bout duration with Novel. Please click here to view a larger version of this figure.
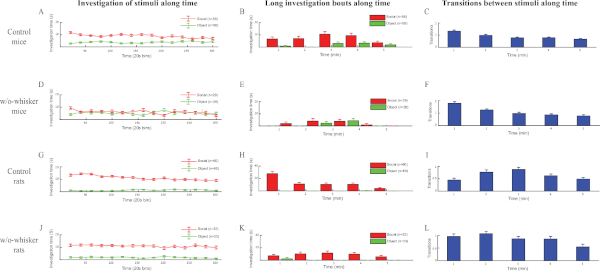
Figure 5: Whisker-trimming of mice and rats differentially impairs social behavior. (A) Plot of mean investigation time (measured in 20 s bins) during the SP test for male C57BL/6J mice (n = 58). (B) Distribution (1 min bins) of mean total time of long (>19 s) investigation bouts along the SP test shown in panel A for social and object stimuli. Note the much longer time of long investigation bouts towards the social stimulus compared to the object. (C) Distribution (1 min bins) of the transition rate along the time of the SP test shown in (A). (D-F) As shown in panels A-C, 29 mice's whiskers were trimmed about 1 week before the test. These animals lost their social preference, as reflected by the (D) general investigation time and (E) distribution of long bouts. They also displayed a higher general rate of transitions (F), suggesting less sustained interactions with the social stimulus. (G-I) As shown in panels A-C, 60 SD rats performed the SP test. Note the different dynamics of the rats' behavior compared to mice, with lower transition rates and longer periods of long investigation bouts in the early phase (2 min) of the test. (J-L) As shown in panels G-H, 33 rats were whisker-trimmed. Note that while the rats did not lose social preference following whisker-trimming as mice did, the dynamics of their social behavior in the first 2 min of the test was changed, showing more transitions and fewer long bouts. Please click here to view a larger version of this figure.
