Cerebroventricular microinjection leads to widespread distribution of the injected solution
Using the workflow and technical scheme of the cerebroventricular microinjection depicted in Figure 1, we administered morpholino oligonucleotides into the adult zebrafish brain. An accurate injection protocol leads to dispersion of the injected liquid throughout the brain and efficient targeting of the cells close to the ventricular surface (Figure 2A). Injections where the glass capillary impales the brain tissue will have a dense fluorescent signal at the point of injection (Figure 2B). If the injected amount is not enough, weak fluorescent signal will be observed (Figure 2C).
Cerebroventricular microinjection can be used for knocking down gene expression at the ventricular region of the adult zebrafish brain
We have shown that vivo morpholinos can penetrate a few cell diameters inside the ventricular surface and hence can efficiently target the ventricular cells (Figures 3A, 3B). Morpholino injection can knockdown gene expression as it will alleviate the production of the corresponding protein (an example is PCNA as shown in Figures 3C, 3D). We have previously demonstrated that knocking down genes using this technique leads to functional consequences during the adult neurogenesis (Figures 3E, 3F) or regeneration response (Figures 3G, 3H). We demonstrated that the efficient knockdown range (more than 50% knockdown) is attained approximately 12 hr after injection of the morpholinos 22. We also observed that efficient knockdown period is until approximately 5 days after injection for the PCNA protein 22. More than 70% knockdown was seen until 3 days after the injection 22. The knockdown curves are dependent on the level of expression of the endogenous proteins and the efficiency of the morpholino molecules; and has to be determined for every gene by the experimenter. In our studies, we did not see that the CVMI procedure compromises the survival of the animal, probably because the administration of the morpholino molecules is local and does not cause a systemic toxicity.
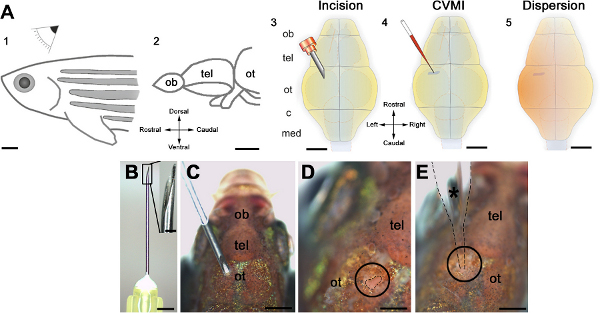
Figure 1. Schematic representation of the cerebroventricular injection technique. (A) The injection is performed from the dorsal of the adult zebrafish (1) at the level of optic tectum (ot, 2). An incision is made over the optic tectal skull plate with a 30-gauge needle (3). Using glass capillaries, the morpholino molecules are injected into the ventricular fluid (4). A thorough dispersion is achieved instantaneously (5). (B) The canula and its barbed-end is shown in high magnification. (C) The orientation of the needle before the incision. (D) The incision is circled and outlined by dashed lines. (E) The injection capillary is located as shown and injection is performed. All panels were adapted from ref. 22. Click here to view larger figure.
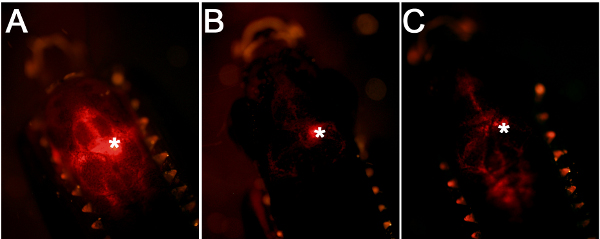
Figure 2. Comparison of injection efficiency and accuracy. (A) A good injection leads to targeting of the cells close to the ventricular surface. (B) If the glass capillary is inserted into the brain tissue, this will generate a centrally located fluorescence signal. (C) If the amount of injection is low, this will lead to weaker fluorescence signal.
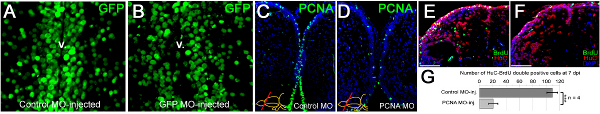
Figure 3. Morpholino penetration, efficiency of gene knockdowns and functional consequences. (A) GFP immunostaining on the telencephalic cross sections of control morpholino-injected Tg(H2A:GFP) transgenic line showing GFP-positive nuclei. (B) GFP immunostaining on the telencephalic cross sections of gfp antisense morpholino-injected Tg(H2A:GFP) transgenic line showing GFP-positive nuclei. Note the difference in the GFP-signal at the ventricular region. v: ventricle. (C) PCNA immunostaining on control morpholino-injected brains showing widespread distribution of PCNA-positive cells. (D) PCNA immunostaining on pcna antisense morpholino-injected brains showing significantly reduced amounts of PCNA-positive cells, showing that CVMI can knockdown endogenous genes. (E) BrdU and HuC/D immunostaining on control morpholino-injected brains to determine newborn neurons after a BrdU pulse-chase experiment described before 22. (F) BrdU and HuC/D immunostaining on pcna antisense morpholino-injected brains to determine newborn neurons after a BrdU pulse-chase experiment described before 22. (G) Knocking-down PCNA leads to significant reduction in generation of newborn neurons, indicating that CVMI can be used to knockdown endogenous genes, which give rise to functional consequences. All panels were adapted from ref. 22 Click here to view larger figure.
