The evaluation of the end yield relies on the level of purity and the quantity of isolated cells and it could be determined by RT-qPCR, cell counting, and protein quantification. The purity of the isolated magnetic cells from the hypothalamus was confirmed by the gene expression analysis of CD11b, C1qa, Gad1, and GFAP with RT-qPCR (Figure 1). CD11b and C1qa are predominantly expressed by microglia in a steady state CNS16 and Gad1 and GFAP serve as a neuronal and astrocytic marker respectively. The total cDNA from one hypothalamus ranged from 55 to 98 ng. It is noteworthy that only the positive fraction exhibited CD11b and C1qa expression, indicating the absence of any potential loss of microglial cells in the negative fraction. Primers are provided in Table 2.
Furthermore, the minor levels of the neuronal and astrocytic markers ensure that the vast majority of the isolated cells are microglia since resident macrophages and monocytes constitute only 10 % of the total immune cell landscape of a healthy brain10,11. These results prove the efficacy of the magnetic selection and pave the way for the application of RT-qPCR to assess the expression levels of all genes of interest in freshly sorted hypothalamic microglial cells. Similarly, an RNAseq analysis can be performed on hypothalamic-sorted microglia to have a comprehensive view of their transcriptome.
The quantity of the isolated cells was determined by cell counting and was found to be ~15,000 cells per hypothalamus (mentioned in protocol step 4.10). Additionally, protein concentration in sorted microglia was quantified, revealing an average concentration of 0.5 µg/µL in CD11b+ cells isolated from two pooled hypothalami (Figure 2). This knowledge allows different assays to be performed depending on a minimum protein threshold. Furthermore, by pooling more hypothalami, a higher concentration can be achieved, expanding the scope of potential analyses. The extracted proteins from hypothalamic microglia facilitate diverse techniques, including AlphaLISA immunoassays and kinase activity assays.
Moreover, the described protocol allows researchers to perform several ex vivo analyses, enabling the exploration of microglial activity and their role in the hypothalamus in both physiological and pathological contexts. The quantification of ROS production represents a crucial aspect of microglial immune response17.Therefore, after separation, microglia can be treated with commercially available reagents that allow the detection of ROS in live cells by exhibiting an increased fluorescent signal upon oxidation. Microglia magnetically isolated from three pooled hypothalami is sufficient to detect ROS by FACS, as shown in Figure 3A, though our results show a quite high variability of cell ROS production among the conditions we analyzed. It is noteworthy that, with the above-mentioned commercial kit (see the Table of Materials), we were also able to assess the viability of sorted Cd11b+ cells, thanks to a cell-impermeant dead cell stain that exhibits an increase in fluorescence upon binding DNA. Specifically, our data show that 99% of cells are alive (Figure 3B). Gating strategies were used to distinguish between alive and dead cells, as well as oxidatively stressed and non-stressed cells, based on their fluorescence properties.
Another main feature of microglial activity is phagocytosis, a dynamic and tightly regulated process that plays a key role in immune surveillance, tissue repair, and the overall homeostasis of the central nervous system18. Remarkably, Cd11b+ cells sorted with magnetic coupled beads can be cultured for 24 h and then incubated with fluorescent latex microbeads at different time points to evaluate, either by FACS or by fluorescence microscopy, alterations in the phagocytic activity under different conditions. Freshly sorted microglia can also be exposed to fluorescent beads or any type of pHrodo-coupled particles (e.g., synaptosomes) directly within the tube, and phagocytosis can be assessed by FACS analysis.
Specifically, we incubated directly in the tube containing sorted cell suspension, microglia isolated from two pooled hypothalami or a whole brain with fluorescent latex beads. Microglia that phagocyted at least one fluorescent bead can be distinguished from non-phagocytic cells by flow cytometry since phagocytic microglia will exhibit an enhanced fluorescence. Negative controls, not incubated with beads, were used for gating. In both our experimental conditions (mice fed with chow vs high-fat diet (HFD)) we observed a very low percentage of phagocytic microglia (Figure 4A). However, this result is shown here to demonstrate the efficacy of this method, which, remarkably, allows also to differentiate between cells that phagocyted 1 beads, 2 beads, 3 beads, and so on (Figure 4B). Finally, all the omics approaches could be applied to isolated hypothalamic microglia. We suggest pooling three hypothalami to have enough cells to perform these assays.
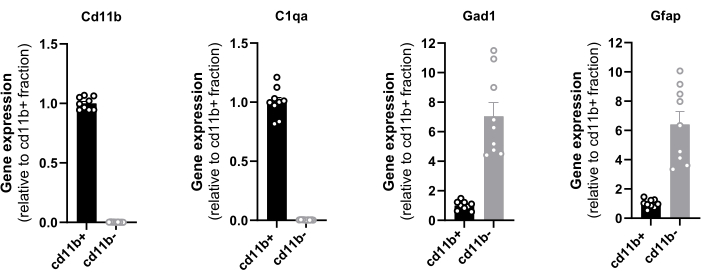
Figure 1: Evaluation of CD11b+ cell purity by RT-qPCR. Quantification in sorted cells of the mRNA expression of the integrin CD11b, complement C1q subcomponent subunit A C1q, glutamic acid decarboxylase Gad1 and glial fibrillary acidic protein GFAP normalized on mRNA levels of housekeeping genes Eef1a1 and Sdha. Please click here to view a larger version of this figure.
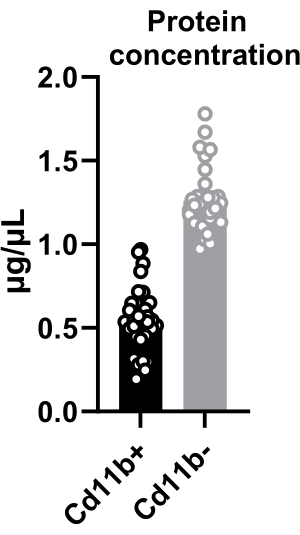
Figure 2: Protein quantification from hypothalamic cells after cell sorting. Quantification of protein concentration in CD11b+ and CD11b– cells. Both CD11b+ and CD11b– cells were lysed in 20 µL of lysis buffer. Please click here to view a larger version of this figure.
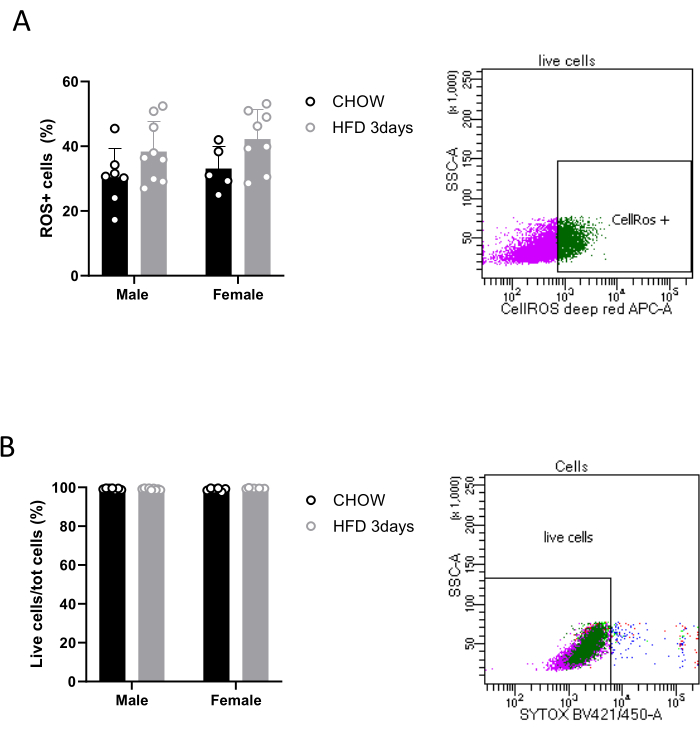
Figure 3: Detection of ROS in microglial live cells. Quantification of (A) oxidatively stressed cells and (B) live cells in a population of sorted Cd11b+ cells, with relative FACS contour plots. Please click here to view a larger version of this figure.
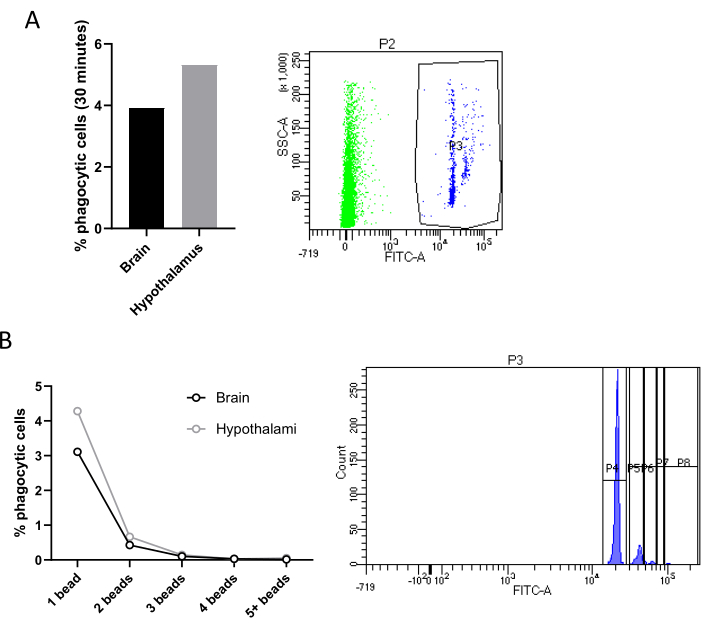
Figure 4: Quantification of microglial phagocytic activity. (A) Quantification of phagocytic cells in Cd11b+ cells sorted from two hypothalami or one brain, with relative FACS contour plot. (B) Percentage of phagocytic cells according to the number of beads phagocyted with relative FACS contour plot. Please click here to view a larger version of this figure.
| Enzyme mix 2 | |||
| Enzyme A | Buffer Y | Enzyme P | |
| 10 µL | 20 µL | 50 µL | |
| D-PBS | Debris Removal Solution | Overlay D-PBS | |
| 15-100 mg structure | 1000 µL | 450 µL | 2 mL |
Table 1: Preparation of the dissociation mixture (volumes refer to a single sample) and the gradient overlayers.
| Genes | Protein | Forward | Reverse | ||||||
| Itgam | Integrin alpha M (CD11b) | CTCATCACTGCTGGCCTATACAA | GCAGCTTCATTCATCATGTCCTT | ||||||
| Sdha | Succinate Dehydrogenase Complex Flavoprotein Subunit A | CCCTCTCATATGTGGACATCAA | CAAGGTCTTGTCGATAACAGGT | ||||||
| C1qa | Complement component 1, q subcomponent, alpha polypeptide | CGAGGTGTGGATCGAAAAGG | GAAGATGCTGTCGGCTTCAGT | ||||||
| Gad1 | glutamate decarboxylase 1 | GACCAATCTCTGTGACTCGCTTAG | CTGGTCAGTGTATCGGAGGTCTT | ||||||
| Gfap | glial fibrillary acidic protein | AGGTCCGCTTCCTGGAACA | GGGCTCCTTGGCTCGAA | ||||||
Table 2: RT-qPCR primer sequences.
| Flow-cytometry cell sorting (FACS) | Magnetically activated cell sorting (MACS) |
| Fluorescent-tagged biochemical antibodies | Magnetic beads conjugated to antibodies |
| Differentiation beyond cell surface markers | Differentiation only according to cell surface markers |
| Substantial initial yield | Modest initial yield |
| Slower than MACS for simultaneous handling of many samples | Faster than FACS for simultaneous handling of many samples |
| Less gentle than MACS | Gentler than FACS |
Table 3: Representation of FACS and MACS characteristics and limitations
