Data processing and analysis
While there are numerous methods to process the raw data from respirometry experiments, it is recommended to use the R package respR28. In keeping with the sharing of the protocols above, which advocate for open science and reproducibility, this package allows data processing and analysis to be shared in an easily reproducible form and has been designed with that in mind. It is a free, open-source platform and probe-system agnostic, and easily installable either from CRAN or GitHub. Full code and examples for respR are maintained and can be found at https://github.com/januarharianto/respR.
The respR package has functions to import, visualize, and perform quality control on respirometry data, and to calculate respiration rates either automatically or from manually chosen regions. It can also adjust rates for background respiration and conversion rates to commonly used output units. The steps to process the data from the micro-respirometry system are detailed below. In this study, the data from the respirometry system was used as an example, but the package also accepts inputs from the majority of commercially available oxygen probe systems as well as generic R data objects. More details about the package, including full documentation and tutorials, can be found on the package website at https://januarharianto.github.io/respR/index.html.
Importing raw data
The raw output file (.txt) is imported. respR recognizes the format and parses it to a generic R data frame that can be used in subsequent functions. However, it is important to note that this is optional; these files and virtually any oxygen time series data can also be imported using base functions (given below) by anyone with a basic knowledge of R.
#load respR
Library(respR)
#Import —
Data <- import_file("file.txt")
#Firesting-Pryo file detected
Inspecting and visualizing the data
A vital part of any data analysis task is to plot and inspect the data to look for obvious anomalies or patterns, or even to just help to understand it. The inspect function is used here (Figure 8A), which checks for issues common to respirometry data such as non-numeric or missing values.
#inspect a single oxygen column
Insp <-inspect(data, time = 3, oxygen = 8, width = 0.2)
This function also plots the oxygen timeseries and calculates a rolling rate (bottom panel) to help elucidate how this rate may change over the course of the experiment. These rolling rate plots help inform about which regions of those rate curves should be extracted. In the case of standard or routine metabolic rates, the desired regions are those where the rate shows stability (e.g., after around timepoint 3,000; Figure 8B).
Here, declining oxygen only becomes detectable after around row 200 in the full timeseries panel. Patterns such as this are very common in respirometry data; there is often an extended period of instability at the start of an experiment as the system stabilizes and specimen becomes acclimated to the experimental conditions. It is recommended that rates should only be extracted from the timeseries after this initial instability, which also highlights the importance of visualizations.
Extract rates
respR has two functions for extracting respiration rates. The first is the calc_rate() function, which allows a rate to be extracted manually by specifying a region of time, row, or oxygen level. This is very common in respirometry analyses, and a perfectly acceptable method of determining a rate as long as selection criteria are decided upon and applied consistently28.
A more robust and objective way is to use the auto_rate() function, which identifies linear regions of the data. These regions are those of consistently sustained respiration rates, assigned automatically using machine learning. This function is also useful for the detection of low signals (as in the samples used in this current study, due to the low biomass at this age). This function allows for the identification of the most linear, minimum, and maximum rates using independent, objective, and statistically robust methods28. The example here identifies a linear region occurring from around timepoints 3,000 to 5,000. It should be noted that multiple linear regions may be identified, but this section is the highest ranked, or most linear, region (Figure 8C).
#Determine most linear (i.e., consistent) rate
rate <-auto_rate(insp)
Background adjustment
Background rates from control experiments can be determined in a similar manner to the above example, and can be used to adjust the specimen rates using the adjust_rate() function (Figure 9A; note that the full analysis is not shown here, only the adjustment). Full examples are detailed on the respR website.
#Adjust rate for background
rate_adj <-adjust_rate(rate, by = bg) #saved bg object
print(rate_adj)
Convert rates
The final step is to convert the rates to the desired output units, using the original units of the raw data, effective volume of the respirometer, and other experimental data, including normalizing to blank measurements (Figure 9B). The output can be an absolute respiration rate, that is, of the entire specimen, or a mass- or surface area-specific rate. The surface area-specific rate was the output used here, which specifically is the absolute rate divided by the surface area of the specimen (Figure 9C).
As discussed above, this system was developed to measure very small specimens. Therefore, we expected low values and potential overlap with blank measurements. Some level of signal is expected within the blanks, and when examined, these values are within the expected range of general experimental noise, possibly due to probe drift, slight temperatures changes, or bubbles on the probes. By design and due to the small specimen size, and therefore small effective volume used, the use of blanks is particularly important here, especially for each run. Representative values have been included here as an example (Figure 10). Given the small specimen size, we recommend the use of the blanks at each run to standardize measurements per run.
These blank values are then used to standardize treatment measurements values. Given corals respire in addition to producing oxygen, the metabolic rate can range from negative to positive values. An example of representative results of the range of respiration values detected from the micro-respiration tool is given here. These results were determined from a successful experiment on single coral juveniles (Figure 10). Overall, respiration was expected to be difficult to detect in this example dataset (by design), given the small size of the samples; this underscores the value of this method in capturing this low signal threshold. These representative results show respiration in the dark at the smallest specimens sizes tested, underscoring the minimum detection threshold of this system. We also measured under two conditions (control and high temperature stress). After standardizing to the blanks measured per run, values ranged from close to zero (control) to a median of ~-5e-5 for the stress treatment. As expected, respiration was low. These results clearly show representative values for blanks, as well as a control versus high temperature comparison for these very small samples.
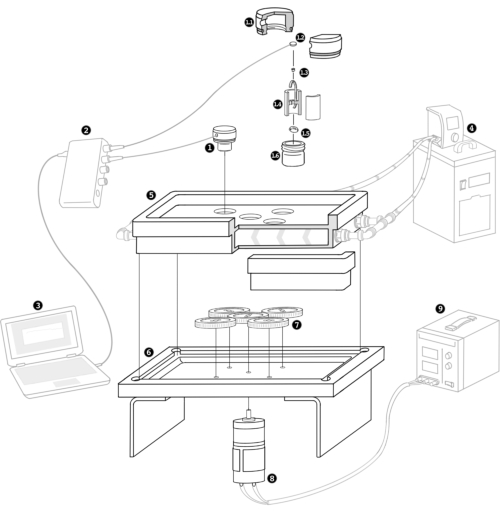
Figure 1: Schematic representation of the new micro-respirometry tool for the physiological characterization of the coral holobiont (coral animal + symbionts) or any small organism (<1 mm). Custom respirometry chambers were made (number 1; 1.1-1.6). These include lids (1.1) with oxygen sensor spots (1.2), and the individual juvenile (1.3) is placed on a flow-through stand (1.4) set atop a magnetic stirrer (1.5), all of which fit within the glass chamber (1.6). The controller (2) is attached to the spot with a fiber optic cable that fits into the lid (1) and connected to the computer (3). The heater/chiller (4) connects to the respirometry plate (5) with the flow-through water (indicated by chevron arrows for direction), which sits on top of the stirrer plate (6) with gears (7), powered by the motor (8) and power supply (9). Please click here to view a larger version of this figure.

Figure 2: Micro-respirometry setup. Multiple options are available, including (A) one respirometry plate, or (B) connected to multiple plates. Please click here to view a larger version of this figure.

Figure 3: Custom built magnetic stirrer plate on top of respirometry plate. Each chamber has (A) a magnetic stirrer gear underneath, (B) powered by a motor, with (C) the respirometry plate connected by tubing to the heater/chiller. Please click here to view a larger version of this figure.

Figure 4: Custom respirometry chamber setup. (A) Components (left to right: lid, glass vial, stand, 1.5 ml tube for scale, and stirrer bar). (B) Individual flow-through stand that the specimen sits within. (C) Top-down view of the flow-through stand. (D) and with the stand placed inside the glass vial with the lid screwed on. Please click here to view a larger version of this figure.

Figure 5: Glass vials placed within the stirrer plate. (A) Custom built stirrer plate with (B) a close-up of the complete glass vial with the lid setup. The juvenile coral can be seen through the lid here (brown dot), on top of the zip-tie, with the fiber optic placed in the lid opening. Please click here to view a larger version of this figure.

Figure 6: Chambers placed upside down, ready for calibration. Please click here to view a larger version of this figure.
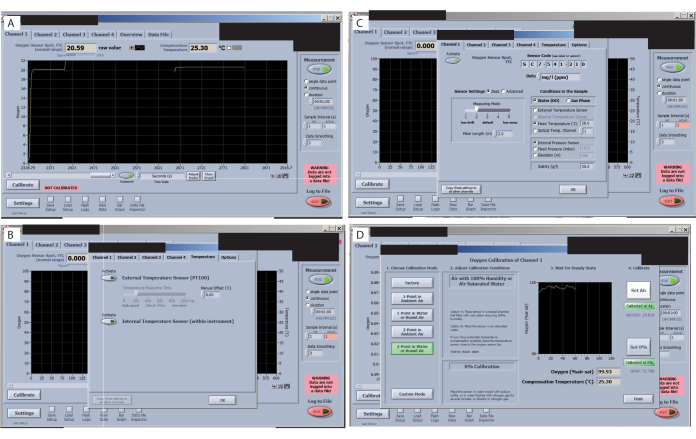
Figure 7: Key steps in the oxygen measurement software. (A) Check the signal of each sensor. An optimal signal for this study and sensors is shown in the oxygen sensor spot FTC (normal range). (B) Check the signal drift. (C) Set and check the treatment temperatures. (D) Set and check the 0% and 100% calibrations. Please click here to view a larger version of this figure.
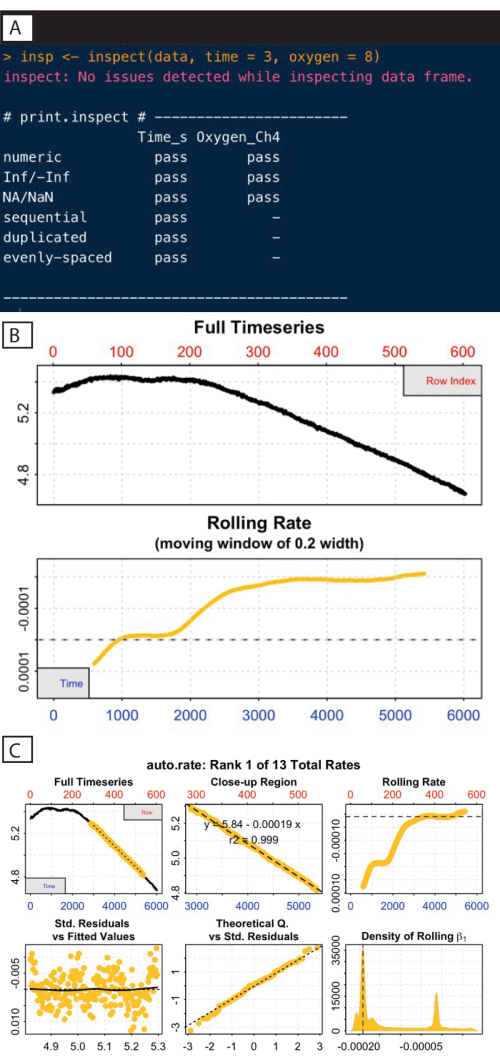
Figure 8: respR analysis output steps I. (A) Inspect the command and output. (B) Check the rate stability. (C) Determine the most linear rate. Please click here to view a larger version of this figure.
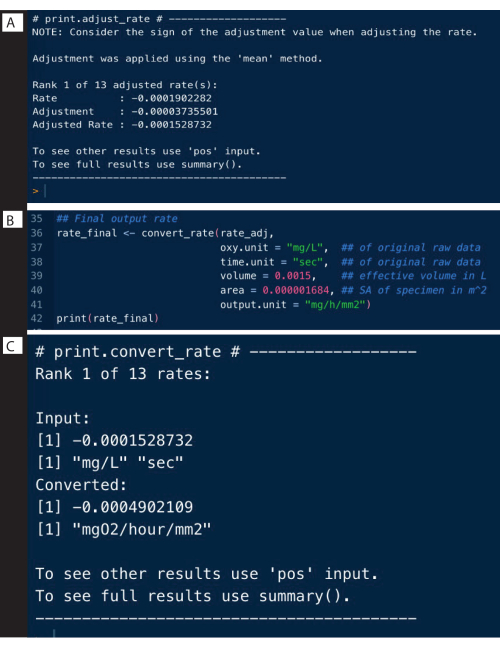
Figure 9: respR analysis output steps II. (A) Adjust the rate for background, (B) Convert and (C) check the rates. Please click here to view a larger version of this figure.
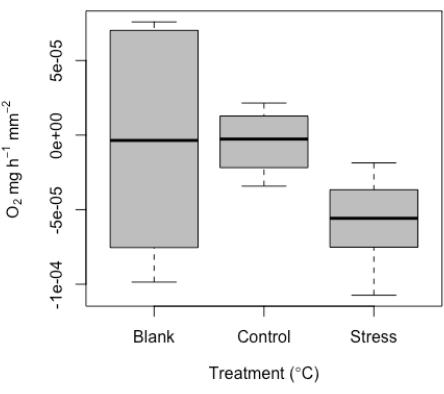
Figure 10: Representative results produced from the micro-respirometry tool. Median respiration (O2 ± standard error) of replicate individual coral juveniles, including blank values as well as respiration of individuals under control and high temperature stress conditions. Please click here to view a larger version of this figure.
Video 1: Top-down view of the respirometry chamber with the juvenile coral inside during a measurement session. Please click here to download this Video.
Table 1: Cost estimates of components of the respirometry apparatus. Please click here to download this Table.
