1. Preparation of the feeding tube
- Punch a hole (~0.3 mm diameter) into the lid of a 2 mL centrifuge tube using an electric winding iron (see Table of Materials). Use such a centrifuge tube to maintain an O. excavata larva and its provision mass.
2. Preparation of pesticide
- Dissolve the technical-grade pesticide (see Table of Materials) in acetone to acquire stock solutions of 1 x 104 µg a.i. mL-1. Then, perform gradient dilutions of the solution to more than five concentrations.
NOTE: Chlorpyrifos at 0.1, 0.2, 0.4, 0.8, 1.6, 3.2, 6.4 µg a.i. mL-1 were used in this study.
3. Preparation of the provisions
- Acquire plastic bee tubes containing provisions (see Table of Materials) and newly hatched larvae of O. excavata from a mass-rearing program.
NOTE: No pesticides were used from 20 days before flowering to the entire flowering period; chemical analysis results showed that commonly used pesticide contents in randomly selected fifty provisions were both below the minimum test levels. - Separate provisions and larvae gently using a soft brush. Select female larvae based on provision size and cell position within the nest9. Then, place uniform-sized provisions and selected female larvae in Petri dishes (60 mm diameter) and set them aside for use.
NOTE: Fifty provisions were randomly selected to analyze the contents of commonly used pesticides: chlorpyrifos, imidacloprid, fendifenuron, phoxim, avermectin. The soft brush parameters are (a) diameter of the brush: 0.3 mm, (b) length of the brush: 2 cm, (c) length of the pen: 18 cm.
4. Provision treatment with pesticide
- Soak the selected evenly sized provisions (from step 3.2) in diluted pesticide (from step 2.1; chlorpyrifos at 0.1, 0.2, 0.4, 0.8, 1.6, 3.2, 6.4 µg a.i. mL-1) for 10 s using a cage. Soak the control check (CK) in 0.2% solvent (acetone in this study).
NOTE: There are three replicates per concentration treatment, and each replicate consisted of 60 provisions. The difference of dosage of each provision can be reduced by selecting evenly sized provisions. - Measure the volume of pesticide solution before and after treating the provisions with the pesticide. Then, calculate the immersed volume of insecticide in each treatment, including 60 mass provisions (Supplementary Table 1). Place the provisions in separate centrifugal tubes with holes (from step 1.1) after air-drying on a sterile worktable.
NOTE: Before the experiment, put the cages containing the provisions into the pesticide solution, and then measure the volume of pesticide solution before and after soaking to eliminate the error. - Transfer female larvae individually to the surface of naturally dried provisions using a soft brush.
NOTE: One larva in one tube.
5. Growth conditions
- Rear the larvae of O. excavata in a growth chamber in the dark, 65%-75% relative humidity, and 25 ± 2 °C16.
6. Examination of the results
- The acute lethal toxicity test
- Measure the mortality of the larvae after placing them onto the treated and the control (CK) provisions for 48 h.
NOTE: The death criteria: when the larvae did not respond to mild touch using a soft brush under black-light lamps22. Black-light lamps were used to simulate the dark growth conditions of larvae and avoid the influence of light on the larvae when checking growth indicators. For eliminating human error, the mortalities with and without removal of larvae from the provisions after 48 h in control groups were also measured. - Weigh 60 provisions before and after 48 h of insect rearing trials to determine the amount of provision consumed by each larva.
- Calculate the dose of pesticide at each concentration consumed by each larva according to the percentage of provision eaten and the pesticide content in each provision.
NOTE: The equation for dose calculation is23:
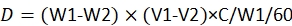
where, D is the consumed dose of pesticide by each larva; W1 is the weight of 60 provisions before infusion of pesticide; W2 is the remaining weight of 60 provisions after 48 h; V1 is the volume of pesticide before immersion for 60 provisions; V2 is the volume of pesticide after immersion for 60 provisions; C is the concentration of the pesticide.
- Measure the mortality of the larvae after placing them onto the treated and the control (CK) provisions for 48 h.
- The sublethal toxicity test
- Weigh the larvae before rearing trials and after 14 days of treatments to determine the larval weight gain.
- Observe O. excavata daily during cocooning under black-light lamps to measure the larval development duration.
- Weigh the remaining portions of provisions after 14 days of feeding on treated and CK provisions to calculate the consumption and the efficiency of conversion of ingested food (ECI)24.
- Examine the number of eclosions by sniping the cocoons using a small scissor when the control bees emerge into adults.
The contents of commonly used pesticides, chlorpyrifos, imidacloprid, fendifenuron, phoxim, avermectin in provisions were less than the limit of quantification (0.01-0.02 mg kg-1) in the control group; these results excluded the influence of pesticide residues on each treatment. The mortality with and without removing larvae from provisions after 48 h in control groups was evaluated; the results showed no significant differences (Table 1), indicating a minor human error.
In the acute lethal toxicity test (Table 2), provisions were soaked in seven diluted pesticide solutions (0.1, 0.2, 0.4, 0.8, 1.6, 3.2, and 6.4 µg a.i. mL-1 chlorpyrifos) and 0.2% acetone (as a control group). A log-probit regression analysis evaluated the median lethal dose (LD50 values) of pesticide to O. excavata according to ingested doses of pesticides (ranging from 0.0001-0.005 µg a.i. mL-1) and corresponding mortality of larvae after 48 h of treatments. The results showed that the LD50 value of chlorpyrifos to the larvae of O. excavata was 0.001 (0.001-0.002) µg a.i. Bee-1.
In the sublethal toxicity test, larval weight gain, developmental duration, eclosion rate, consumption, and ECI of O. excavata were evaluated under the soaking concentrations of 0.1, 0.2, 0.4, and 0.8 µg a.i. mL-1 of chlorpyrifos. An analysis of covariance (ANCOVA) was used to determine treatment-related changes in the development (except eclosion rate) and food utilization of O. excavata. In contrast, initial provision mass was used as a covariate. As the dose increased, the index values of larval weight gain, consumption, and ECI decreased for treatments, with the lowest values relative to the control observed in 0.013 µg a.i. bee-1 chlorpyrifos. Conversely, the most extended larval developmental duration was observed in 0.016 µg a.i. bee-1 chlorpyrifos compared to the control treatment (Figure 1).
Chlorpyrifos's impacts on eclosion rate were evaluated using one-way analysis of variance (ANOVA) and Tukey's least significant difference (LSD) test. Pearson's correlation was also conducted to analyze the relationship between ingested dosages of chlorpyrifos and the eclosion rate of O. excavata. Here, the results of this analysis showed that a significant negative linear relationship exists for the treatments (R2 = 0.82, P = 0.03). The eclosion rate was considerably lower when the ingested dosages exceeded 0.002 µg a.i. bee-1 than those in the control treatment (Figure 2).
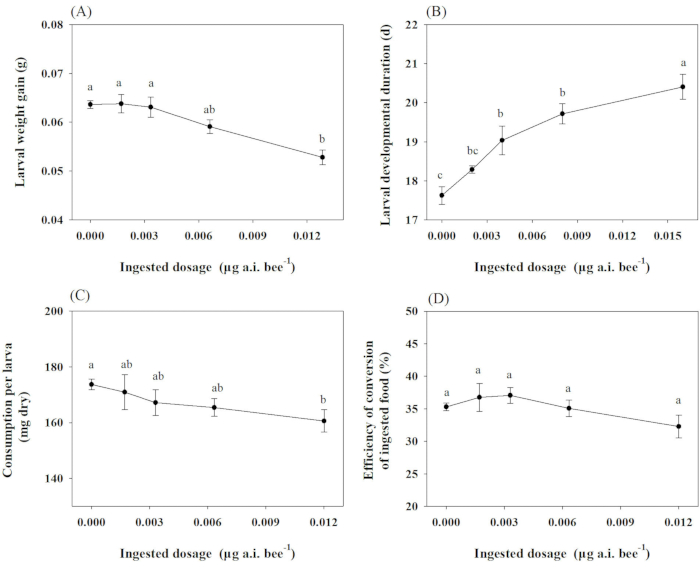
Figure 1: Effect of chlorpyrifos on the growth, development, and feeding of O. excavate. (A), (C), (D): after 14 days of treatment; (B): before cocooning of O. excavate. Different lowercase letters indicate significant differences between treatments at P < 0.05. The numbers for each data point average with SD. Please click here to view a larger version of this figure.
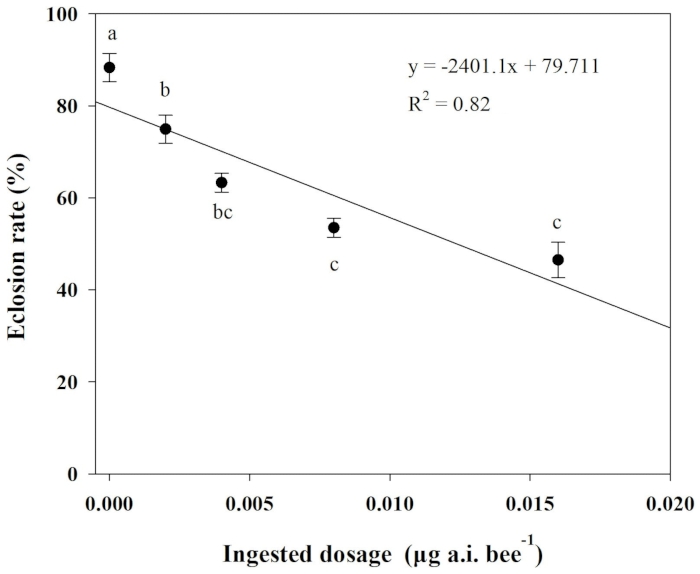
Figure 2: Relationship between ingested dosages of chlorpyrifos and eclosion rate of O. excavate. Different lowercase letters indicate significant differences between the treatments at P < 0.05; The numbers for each data point average with SD. Please click here to view a larger version of this figure.
| Treatments | Mortality | |
| Repetition | Mean | |
| With removal of larvae | 11.91% | 9.67% a |
| 7.63% | ||
| 9.46% | ||
| Without removal of larvae | 6.88% | 8.28% a |
| 7.37% | ||
| 10.59% | ||
Table 1: The mortality with and without removing larvae from provisions after 48 h in control groups. The same lowercase letters indicate no significant differences between treatments at P < 0.05.
| Insecticide | Slope ± SE | Df | χ2 (P) | LD50 (95% CI) (μg a.i. bee−1) |
LD90 (95% CI) (μg a.i. bee−1) |
| Chlorpyrifos | y=3.23+0.30x | 5 | 5.38 (0.37) | 0.001 (0.001-0.002) | 0.02 (0.012-0.038) |
Table 2: Toxicity of Chlorpyrifos to Osmia excavata after 48 h of treatments. SE – standard error; Df – degree of freedom; χ2– values of Chi-square; CI – confidential interval.
Supplementary Table 1: The immersed volume of insecticide in each treatment, including 60 mass provisions. Please click here to download of this Table.
