Purification of Prominin-1+ Stem Cells from Postnatal Mouse Cerebellum
Özet
Demonstrated here is an efficient and cost-effective method to purify, culture, and differentiate white matter stem cells from postnatal mouse cerebellum.
Abstract
Most cerebellar neurons arise from two embryonic stem niches: a rhombic lip niche, which generates all the cerebellar excitatory glutamatergic neurons, and a ventricular zone niche, which generates the inhibitory GABAergic Purkinje cells, which are neurons that constitute the deep cerebellar nuclei and Bergman glia. Recently, a third stem cell niche has been described that arises as a secondary germinal zone from the ventricular zone niche. The cells of this niche are defined by the cell surface marker prominin-1 and are localized to the developing white matter of the postnatal cerebellum. This niche accounts for the late born molecular layer GABAergic interneurons along with postnatally generated cerebellar astrocytes. In addition to their developmental role, this niche is gaining translational importance in regards to its involvement in neurodegeneration and tumorigenesis. The biology of these cells has been difficult to decipher because of a lack of efficient techniques for their purification. Demonstrated here are efficient methods to purify, culture, and differentiate these postnatal cerebellar stem cells.
Introduction
The cerebellum has been long recognized as a major neuronal circuit coordinating voluntary movement1. It receives input from the wide swathes of the neuroaxis, which includes proprioceptive information from the periphery, so as to fine tune motor output and coordinate motion. More recently, it has also been implicated in regulating cognition and emotions by potentially using similar information processing networks2,3,4.
The adult cerebellum is composed of an outer cerebellar cortex and inner white matter. Interspersed within these structures are deep intracerebellar nuclei. Similar to the rest of the nervous system, the development of the cerebellum is driven by the proliferation of multipotent progenitor cells (stem cells) that migrate and differentiate to yield this well-organized structure. In early development (E10.5–E13.5), a ventricular stem niche around the developing fourth ventricle generates GABAergic neurons (i.e., Purkinje cells, Lugaro cells, Golgi cells) along with Bergmann glia5,6,7,8.
Later in development (postnatal week one), a second stem cell niche in the rhombic lip generates MATH1- and Nestin-expressing progenitors that give rise to excitatory granule neurons9,10,11,12. Recently a third stem cell niche has been described13. These cells express prominin-1 (also known as CD133), a membrane-spanning glycoprotein that defines a subset of stem cells in the intestine and hematopoietic systems14,15,16. In vivo fate mapping shows that these stem cells generate key molecular layer interneurons (i.e., basket cells and stellate cells), along with astrocytes, during the first three postnatal weeks. In the past, it has been difficult to study these cells in vitro because prior methods have required costly and time-consuming techniques (i.e., fluorescence-activated cell sorting [FACS]) that are dependent on prominin-1 staining12,13,17. This protocol describes an immunomagnetic-based method for the isolation of these stem cells that can then be readily cultured and differentiated.
Protocol
All animal experiments were performed in compliance with the NIH’s Guide for the Care and Use of Laboratory Animals (2011) and were approved by the Northwestern University IACUC (Protocol IS00011368).
1. Preparation of Solutions
- Prepare tissue dissociation solution made from sterile phenol red-containing Dulbecco’s phosphate-buffered saline (DPBS) with papain (100 U/ml), cysteine (0.2 mg/ml) and DNase (250 U/ml).
- To prepare DNase solution dilute 100 mg of the lyophilized powder of DNase I (one bottle) in 50 mL of H2O. Mix well and filter the stock solution. Prepare 10 mL stock aliquots. Divide one stock tube into 0.5 mL single use aliquots. Store these single use aliquots at -80 °C.
- Prepare magnetic separation reagents by preparing magnetic column buffer X: 0.5% bovine serum albumin (BSA) and 2 mM EDTA solution.
- To prepare neurosphere media, use neurobasal medium containing penicillin/streptomycin with L-glutamine and supplement with 2% B27, 20 ng/mL human recombinant epidermal growth factor (EGF), and 20 ng/mL human recombinant basic fibroblast growth factor (bFGF).
- To prepare differentiation media, use neurobasal medium and supplement with 10 ng/mL differentiated factor platelet-derived growth factor (PDGF-AA) or 10 ng/mL leukemia inhibitory factor (LIF) and 2% B27.
- Use ultra-low attachment 12 well (3.5 cm2) and 6 well (9.6 cm2) culture plates.
2. Dissection of Cerebellum
- Anesthetize mouse pups (P3-P7) with isoflurane and decapitate using surgical scissors.
- Spray the separated head of the pups with 70% ethanol.
- Transfer each head into an empty sterile 10 cm culture dish. Separate the skin using the microdissection scissors, then remove the skull by running the scissors sagittal along the midline.
- Using #7 forceps, peel off the skull bones starting caudally from the brainstem. Carefully lift the brain using a spatula, keeping the cerebellum intact, and transfer the brain to a fresh sterile 10.0 cm dish containing 15 mL of ice-cold Hanks' Balanced Salt solution (HBSS) solution.
- Place the dish containing the brain under a dissection microscope. Using fine #5 forceps, remove the meninges and large blood vessels from the cerebellum and separate the cerebellum from the brainstem using the spatula.
- Transfer the cerebellum into a 15 mL centrifuge tube containing 5.0 mL of ice-cold HBSS solution. Wash the cerebellum by rinsing and decanting 3x with 5.0 mL of HBSS.
NOTE: Each cerebellum should be placed in its own tube for further processing.
3. Cell Suspension Preparation
- After the last rinse, add 5 mL of papain-based tissue dissociation solution (based on previous work13,18) that has been pre-warmed to 37 °C. Incubate the tissue for 15 min at 37 °C in a water bath. Slowly mix the content by inverting the tube up and down 3x–5x every 3 min either using a nutating mixer or by hand.
- Prepare a wide diameter (regular glass pipette) and narrow diameter Pasteur pipette (fire-polished by heating over a Bunsen burner) as described previously19.
- Wash the tissue 3x with 5 mL of HBSS solution, avoiding loss of the tissue while decanting by hand.
- Remove the last HBSS wash and add 5 mL of DPBS solution containing 250 μL of DNase solution to the tissue. Dissociate the tissue by triturating 10x–15x using the wide diameter Pasteur pipette. Perform this step gently to avoid the formation of bubbles.
- Then incubate the slurry for an additional 10 min at 37 °C in the water bath, mixing by inverting and straightening the tube.
- Use the reduced diameter Pasteur pipette to further triturate the tissue slurry 10x. If large pieces of tissue remain, press the tissue pieces against the bottom of the tube with the tip of the pipette gently and continue pipetting until the cells reach a fine suspension.
- Incubate the tissue at 37 °C for an additional 10 min, repeating the previous mixing steps.
4. Immunolabeling of Stem Cells
- Place the centrifuge tubes on ice and use ice-cold solutions for the next steps. Strain the dissociated cells through a 40 μm cell strainer into a 50 mL centrifuge tube. Top the filter with 10.0 mL of HBSS solution to ensure that cells pass through the mesh in this additional solution.
- Transfer the filtered cells into a fresh 15 mL centrifuge tube and centrifuge the cell suspension at 300 x g for 10 min at 4 °C. Aspirate and discard the supernatant completely by carefully using a vacuum aspirator.
- Resuspend the pellet in 160 μL of magnetic column buffer. To obtain the single-cell suspension before magnetic labeling, pass cells through a 30 μm nylon mesh to remove cell clumps, which can otherwise clog the column.
- Add 40 μL of anti-prominin-1 microbeads to each 15 mL tube, mix, and incubate in a refrigerator for 15 min in order for the antibody to bind to prominin-1-expressing cells.
- Wash the cells by adding 1.0–2.0 mL of column buffer X and centrifuge at 300 x g for 10 min. Aspirate the supernatant completely.
- Resuspend the pellet in 1.0 mL of column buffer X.
NOTE: If small clumps are seen after the resuspension of pellet with 1.0 mL of column buffer X, they should be removed carefully using a tip of the Pasteur pipette, otherwise these clumps can block the magnetic column during cell sorting.
5. Magnetic Column Preparation, Cell Sorting, and Plating
NOTE: The magnetic separation from different genotype conditions (disease vs. control) must be performed at same time, since any delay may affect the neurosphere morphology.
- Prepare the magnetic columns by placing them on the magnetic stand exposed to the magnetic field. Rinse the column once with 500 μL of buffer X by applying buffer that drips into a centrifuge tube to be discarded.
NOTE: Prepare fresh magnetic column buffer X for each experiment; if not, then stem cell yield will be low. - Apply the labeled cell suspension onto the column. Collect the flowthrough containing unlabeled cells that mainly consist of cerebellar neuronal/glial mixed cells into fresh 15 mL tubes (Figure 1A).
- Wash the column 3x with 500 μL of buffer X (each wash takes around 2–4 min).
NOTE: Cerebellar neuronal/glial mixed culture enriched with cerebellar granular neurons can serve as a useful byproduct of this purification step. - Remove the column from the magnetic field and place in a 1.5 mL tube. Add 1.0 mL of culture medium (neurosphere medium) to the column and push the plunger into the column to flush out the cells tagged with prominin-1 beads into a fresh 1.5 mL falcon tube.
- To enhance the purity of prominin-1 labeled cells, pass the eluted cells over a second column following steps 5.1–5.4.
- Count the cells with a hemocytometer. The typical yield is 107 cells per cerebellum. Plate the cells onto ultra-low attachment plates (6 or 12 well, based on the density required for downstream experiments).
6. Passaging of Neurospheres and Differentiation
- Plate the prominin-1-labeled stem cells on ultra-low attachment 12 well plates in neurosphere medium (5,000 cells/well).
- After 7–10 days, the cells divide to yield ball shaped floating neurospheres (primary neurospheres).
NOTE: In this experiment, secondary neurospheres that expand in numbers are generated for further use. Primary neurosphere populations are not used for these experiments, since they may contain contaminating cells that do not have stem cell properties and can clump together with neurospheres. - For passaging, transfer the primary neurospheres along with the culture media using 1.0 mL pipette tips to a 15 mL sterile centrifuge tube. Pellet the neurospheres by centrifugation at 300 x g for 5 min and discard the supernatant.
- Resuspend the pellet in 5 mL of tissue dissociation media that contains papain or 0.05% trypsin solution. Incubate at 37 °C for 10 min.
- Centrifuge the cell suspension at 300 x g for 5 min. Resuspend the cells in 5 mL of neurosphere medium and dissociate the cells mechanically using a plastic Pasteur pipette and slowly pipetting up and down 10x.
- Plate the cells (again) in neurosphere media as described earlier. After 7–10 days in culture, the plate should be enriched with secondary neurospheres.
NOTE: These cultures can be passaged up to 8x with good efficiency, after which neurosphere size tends to be smaller and suggestive of a decrease in proliferation. - Count the cells on the hemocytometer and plate the optimal number for future experiments on poly-D-lysine coated plates (6 or 12 wells).
- To differentiate the stem cells, collect neurospheres from the second to eighth passage by centrifugation as described earlier, except add differentiation medium to the pellet.
NOTE: After 7 days in vitro, the stem cells differentiate into neurons, astrocytes, and oligodendrocytes, as demonstrated by staining with the neuronal marker β-III tubulin, astrocytic marker GFAP, and oligodendrocyte maker O4.
Representative Results
Prominin-1-positive postnatal cerebellar stem cells formed neurospheres in neurosphere medium rich in growth factors (EGF and bFGF). These neurospheres were positive for prominin-1-staining, the marker used for isolation, and also as a stain for other stem cell markers such as Nestin and GFAP13 (Figure 1). The stem cell marker expression was maintained throughout culture and for up to at least eight passages20. Upon withdrawal of growth factors and in the presence of LIF and PDGF-AA (which are factors that support neuronal and glial differentiation21,22), the neurospheres differentiated into neuronal and glial lineages (Figure 2).
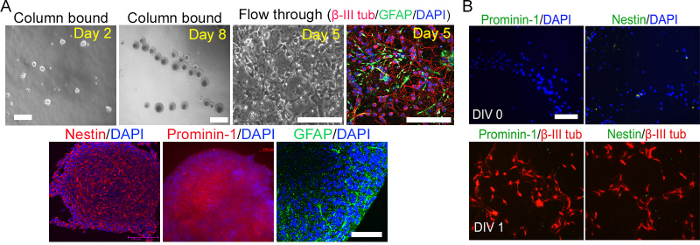
Figure 1: Isolation of prominin-1 stem cells from postnatal cerebellum. (A) Cerebellar stem cells were isolated using immunomagnetic prominin-1 beads. Top panel: Purified stem cells (column-bound) formed neurospheres with extensive proliferation and self-renewal properties. The cells unbound to the column (flowthrough) were unable to form neurospheres; instead, they became cerebellar neuronal/glial mixed cells (β-III tubulin/GFAP). Bottom panel: the neurospheres formed from prominin-1+ cells expressing stem cell-specific markers: Nestin, prominin-1, and GFAP. (B) Cells in the flowthrough stained negative for stem cell markers prominin-1 and nestin and positive for neuronal marker β-III tubulin. Please click here to view a larger version of this figure.
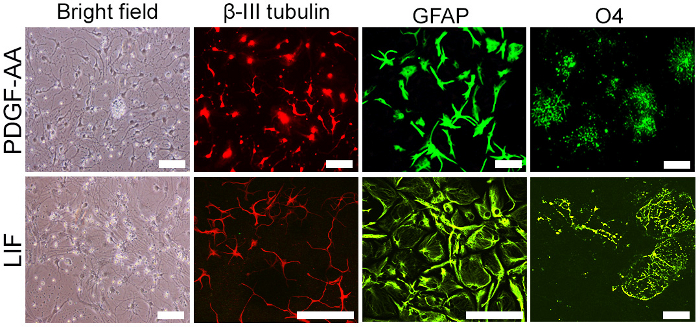
Figure 2: Differentiation of prominin-1 positive neurospheres. In the presence of differentiation factors (PDGF-AA or LIF), prominin-1-positive neurospheres differentiated into neurons (β-III tubulin), astrocytes (GFAP), and oligodendrocytes (O4). Please click here to view a larger version of this figure.
Discussion
Prominin-1-expressing cerebellar stem cells reside in the prospective white matter during the first 3 weeks of postnatal life. Their proliferation is tightly controlled by the sonic hedgehog pathway supported by Purkinje cells17. These stem cells/progenitors contribute to later-born GABAergic interneurons called basket cells and stellate cells. These interneurons reside in the molecular layer, where they synapse onto Purkinje cells and sculpt PC topography and function via GABAergic inhibition13,17,23. Besides forming interneurons, this stem cell population also generates all postnatally derived cerebellar astrocytes17,24.
This protocol describes an easy and cost-effective method to purify prominin-1/CD133 stem cells from the postnatal mouse cerebellum. Stem cells must be cultured in ultra-low attachment plates. Culturing these stem cells in normal plates may cause the neurospheres to attach to the surface and lead to low stem cell proliferation and differentiation. Here, the yield of 200–300 neurospheres per 5,000 cells corresponds to a stem cell yield of around 1 x 107 cells from a single cerebellum. This is comparable to what has been described for an FACS-based strategy that is 10x more expensive. Moreover, FACS equipment requires an expensive set-up and highly trained personnel, and is not readily available.
These stem cells are also gaining increasing translational significance in research on cancer as well as neurodevelopmental and neurodegenerative disorders25,26,27. Uncontrolled proliferation of these stem cells in early life leads to medulloblastoma28, while research from our own lab suggests that their abnormal proliferation and differentiation can contribute to later cerebellar degeneration in the genetic disease spinocerebellar ataxia type 120. These new protocols will be valuable for studying these cells and provide novel insight into their roles in health and disease. These methods may also lead to advances in regenerative therapies after stroke or trauma and other insults to the brain that would warrant neuroregeneration. It is conceivable that these techniques can be generalized to extract prominin-1-expressing stem cells from other tissues, such as intestine and bone marrow, where they are also expressed15,29.
Açıklamalar
The authors have nothing to disclose.
Acknowledgements
We thank the Opal lab members for their suggestions. This work was supported by NIH grants 1RO1 NS062051 and 1RO1NS08251 (Opal P)
Materials
| 0.05%Trypsin | Thermo Fisher Scientific | 25300054 | 0.05% |
| 2% B27 | Gibco; Thermo Fisher Scientific | 17504001 | |
| 2mM EDTA solution | Corning | 46-034-CI | |
| Anti- Prominin-1 microbeads | Miltenyi Biote | 130-092-333 | |
| bovine serum albumin | Sigma | A9418 | |
| Column MultiStand | Miltenyi Biotec | 130-042-303 | |
| culture plates ultra – low attachment | Corning | 3473 | |
| cysteine | Sigma | C7880 | |
| DNase | Sigma | D4513-1VL | 250 U/ml |
| Dulbecco’s Phosphate Buffer Saline | Thermo Fisher Scientific | 14040141 | |
| Hank's balanced salt solution-HBSS | Gibco | 14025-092 | |
| Human recombinant Basic Fibroblast Growth Factor | Promega | G507A | 20 ng/ml |
| Human recombinant Epidermal Growth Factor | Promega | G502A | 20 ng/ml |
| Leukemia Inhibitory Factor | Sigma | L5158 | |
| l-glutamine | Gibco | 25030081 | |
| Microscopy | Lieca TCS SP5 confocal microscopes | ||
| MiniMACS separator | Miltenyi Biotec | 130-042-102 | |
| mouse anti-Prominin-1 | Affymetrix eBioscience | 14-1331 | 1 in 100 |
| Nestin | Abcam | ab27952 | 1 in 200 |
| Neurobasal medium | Thermo Fisher | 25030081 | |
| O4 | Millopore | MAB345 | |
| Papain | Worthington | LS003126 | (100 U/ml) |
| Platelet- Derived Growth Factor | Sigma | H8291 | 10 ng/ml |
| Poly-D-Lysine | Sigma | P6407 | |
| rabbit anti-tubulin, b-III | Sigma | T2200 | 1 in 500 |
| Rabit anti-GFAP | Dako | Z0334 | 1 in 500 |
| Separation columns-MS columns | Miltenyi Biotec | 130-042-201 | |
| Sterile cell strainer | Fisher Scientific | 22363547 | 40um |
Referanslar
- Glickstein, M., Strata, P., Voogd, J. Cerebellum: history. Nörobilim. 162, 549-559 (2009).
- Carta, I., Chen, C. H., Schott, A. L., Dorizan, S., Khodakhah, K. Cerebellar modulation of the reward circuitry and social behavior. Science. 363, (2019).
- Sathyanesan, A., et al. Emerging connections between cerebellar development, behaviour and complex brain disorders. Nature Reviews Neuroscience. 20, 298-313 (2019).
- Wagner, M. J., Kim, T. H., Savall, J., Schnitzer, M. J., Luo, L. Cerebellar granule cells encode the expectation of reward. Nature. 544, 96-100 (2017).
- Araujo, A. P. B., Carpi-Santos, R., Gomes, F. C. A. The Role of Astrocytes in the Development of the Cerebellum. Cerebellum. , (2019).
- Seto, Y., et al. Temporal identity transition from Purkinje cell progenitors to GABAergic interneuron progenitors in the cerebellum. Nature Communication. 5, 3337 (2014).
- Marzban, H., et al. Cellular commitment in the developing cerebellum. Frontiers in Cell Neurosciences. 8, 450 (2014).
- Koziol, L. F., et al. Consensus paper: the cerebellum’s role in movement and cognition. Cerebellum. 13, 151-177 (2014).
- Ben-Arie, N., et al. Math1 is essential for genesis of cerebellar granule neurons. Nature. 390, 169-172 (1997).
- Machold, R., Fishell, G. Math1 is expressed in temporally discrete pools of cerebellar rhombic-lip neural progenitors. Neuron. 48, 17-24 (2005).
- Wang, V. Y., Rose, M. F., Zoghbi, H. Y. Math1 expression redefines the rhombic lip derivatives and reveals novel lineages within the brainstem and cerebellum. Neuron. 48, 31-43 (2005).
- Li, P., et al. A population of Nestin-expressing progenitors in the cerebellum exhibits increased tumorigenicity. Nature Neurosciences. 16, 1737-1744 (2013).
- Lee, A., et al. Isolation of neural stem cells from the postnatal cerebellum. Nature Neurosciences. 8, 723-729 (2005).
- Toren, A., et al. CD133-positive hematopoietic stem cell “stemness” genes contain many genes mutated or abnormally expressed in leukemia. Stem Cells. 23, 1142-1153 (2005).
- Zhu, L., et al. Prominin 1 marks intestinal stem cells that are susceptible to neoplastic transformation. Nature. 457, 603-607 (2009).
- Man, S. M., et al. Critical Role for the DNA Sensor AIM2 in Stem Cell Proliferation and Cancer. Cell. 162, 45-58 (2015).
- Fleming, J. T., et al. The Purkinje neuron acts as a central regulator of spatially and functionally distinct cerebellar precursors. Developmental Cell. 27, 278-292 (2013).
- Panchision, D. M., et al. Optimized flow cytometric analysis of central nervous system tissue reveals novel functional relationships among cells expressing CD133, CD15, and CD24. Stem Cells. 25, 1560-1570 (2007).
- Beaudoin, G. M., et al. Culturing pyramidal neurons from the early postnatal mouse hippocampus and cortex. Nature Protocols. 7, 1741-1754 (2012).
- Edamakanti, C. R., Do, J., Didonna, A., Martina, M., Opal, P. Mutant ataxin1 disrupts cerebellar development in spinocerebellar ataxia type 1. Journal of Clinical Investigation. 128, 2252-2265 (2018).
- Erlandsson, A., Enarsson, M., Forsberg-Nilsson, K. Immature neurons from CNS stem cells proliferate in response to platelet-derived growth factor. Journal of Neurosciences. 21, 3483-3491 (2001).
- Galli, R., Pagano, S. F., Gritti, A., Vescovi, A. L. Regulation of neuronal differentiation in human CNS stem cell progeny by leukemia inhibitory factor. Developmental Neurosciences. 22, 86-95 (2000).
- Silbereis, J., Cheng, E., Ganat, Y. M., Ment, L. R., Vaccarino, F. M. Precursors with Glial Fibrillary Acidic Protein Promoter Activity Transiently Generate GABA Interneurons in the Postnatal Cerebellum. Stem Cells. 27, 1152-1163 (2009).
- Parmigiani, E., et al. Heterogeneity and Bipotency of Astroglial-Like Cerebellar Progenitors along the Interneuron and Glial Lineages. Journal of Neurosciences. 35, 7388-7402 (2015).
- Wojcinski, A., et al. Cerebellar granule cell replenishment postinjury by adaptive reprogramming of Nestin(+) progenitors. Nature Neurosciences. 20, 1361-1370 (2017).
- Yang, Z., Joyner, A. L. YAP1 is involved in replenishment of granule cell precursors following injury to the neonatal cerebellum. Gelişim Biyolojisi. 1606 (19), 30207 (2019).
- Wang, S. S., Kloth, A. D., Badura, A. The cerebellum, sensitive periods, and autism. Neuron. 83, 518-532 (2014).
- Eberhart, C. G. Three down and one to go: modeling medulloblastoma subgroups. Cancer Cell. 21, 137-138 (2012).
- Takahashi, M., et al. CD133 is a positive marker for a distinct class of primitive human cord blood-derived CD34-negative hematopoietic stem cells. Leukemia. 28, 1308-1315 (2014).
