To determine the efficiency of the dendrimer-mediated siRNA delivery, we resected the apex of the ventricle of the zebrafish heart, then injected about 10 µL of dendrimer only (mock group), Cy5-siRNA only (naked group), or f-PAMAM-PEG-R9 dendrimer-encapsulated Cy5-siRNA (Cy5-siRNA group) intrapleurally, respectively (Figure 2A-B). The fluorescence signal was detectable in the hearts injected with dendrimer-encapsulated Cy5-siRNA at 3, 24, and 48 hpi (hours post-injection), while it was hardly detectable in hearts from the mock and naked groups at 48 hpi (Figure 2C-D), suggesting that the f-PAMAM-PEG-R9 dendrimer effectively facilitates the delivery of siRNAs into the adult zebrafish heart and is stable for at least two days.
To investigate the effect of dendrimer-encapsulated siRNA on gene silencing, we chose aldh1a2 (retinoic acid-synthesizing enzyme) as a target gene, which has been reported to be required for heart regeneration after ventricular resection44. As shown in Figure 3A, the fish were allowed to recover for one day after ventricular resection and then microinjected with dendrimer-encapsulated siRNA. We found that the aldh1a2 mRNA expression level decreased in the hearts treated with f-PAMAM-PEG-R9 dendrimer-encapsulated siAldh1a2 compared with that of dendrimer-encapsulated scrambled siRNA (siNC) at 2 dpa (Figure 3B), demonstrating that f-PAMAM-PEG-R9 dendrimer-mediated siRNAs achieve gene-specific silencing in adult zebrafish heart.
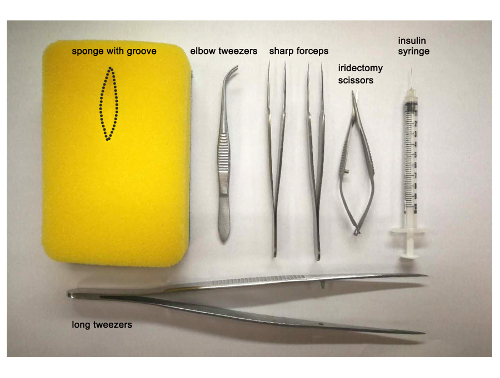
Figure 1: Heart surgery and thoracic injection instruments. Photograph of the instruments used for ventricular resection in the adult zebrafish heart: sponge with a groove, elbow tweezers, sharp forceps, iridectomy scissors, long tweezers, and the insulin syringe used for dendrimer-encapsulated siRNA delivery. Please click here to view a larger version of this figure.
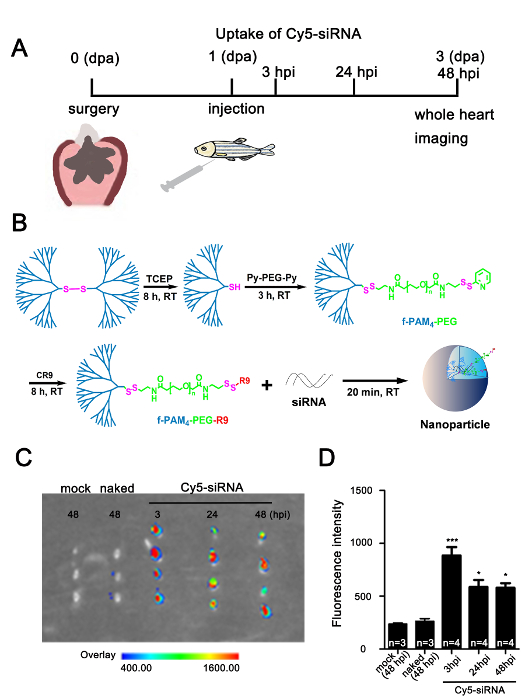
Figure 2: Effective f-PAMAM-PEG-R9 dendrimer-assisted delivery of siRNA into the injured adult zebrafish heart. (A) Scheme for investigating the uptake efficiency of nanoparticle-encapsulated Cy5-siRNA in adult zebrafish heart after ventricular resection (dpa: days post-amputation; hpi: hours post-injection). (B) Synthesis of f-PAMAM-PEG-R9 dendrimer and preparation of dendrimer-encapsulated Cy5-siRNA. (C) Cy5 fluorescence imaging of hearts injected with dendrimers only (mock), Cy5-siRNA only without dendrimer encapsulation (naked), and dendrimer-encapsulated fluorescence labeled Cy5-siRNA (Cy5-siRNA) detected by the in vivo imaging system, showing that fluorescence signals were nearly undetectable in the mock and naked groups at 48 hpi, while strong signals retained in the dendrimer-encapsulated Cy5-siRNA groups from 3 to 48 hpi. The arbitrary scale of fluorescence signals shows from weak (blue) to strong (red). (D) Quantification of Cy5-siRNA fluorescence signals in hearts assessed by the in vivo imaging system as in panel C (*P <0.05, ***P <0.001; data are mean ± s.e.m.; one-way analysis of variance followed by Bonferroni's multiple comparison tests; n = 3-4 hearts). Please click here to view a larger version of this figure.
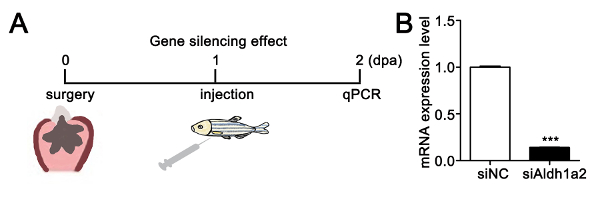
Figure 3: Efficient siRNA silencing of aldh1a2 in the injured adult zebrafish heart. (A) Scheme for investigating the gene-silencing effect of aldh1a2 siRNA. (B) qPCR reveals that aldh1a2 mRNA decreased in nanoparticle-encapsulated siAldh1a2 hearts compared with nanoparticle-encapsulated scrambled siRNA hearts (siNC). The aldh1a2 mRNA expression level was normalized by GAPDH (***P <0.001; data are mean ± s.e.m. with paired Student's t-test). Please click here to view a larger version of this figure.
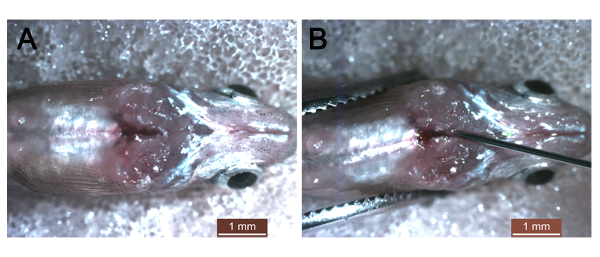
Supplementary Figure 1: Images of thoracic cavity injection. (A) The thoracic cavity in adult fish at 1 day post amputation. (B) Injection of nanoparticle-encapsulated siRNA into the thoracic cavity. Scale bars: 1 mm. Please click here to view a larger version of this figure.
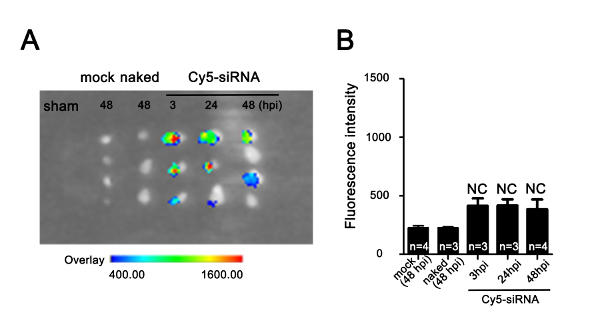
Supplementary Figure 2: Uptake efficiency of nanoparticle-encapsulated Cy5-siRNA in uninjured adult zebrafish heart. (A) Cy5 fluorescence imaging of hearts injected with dendrimers only (mock), naked Cy5-siRNA without dendrimer encapsulation (naked), and dendrimer-encapsulated Cy5-siRNA under the in vivo imaging system. The arbitrary scale of fluorescence signals is from weak (blue) to strong (red). (B) Quantification of Cy5-siRNA fluorescence signals in hearts measured by the in vivo imaging system as in panel A (NC: no significant different; data are mean ± s.e.m.; one-way analysis of variance followed by the Bonferroni's multiple comparison test; n = 3-4 hearts). Please click here to view a larger version of this figure.
