In a representative clonal analysis of SMCs in embryos mutant for Eln (the gene encoding the extracellular matrix protein elastin), Eln(+/-), Acta2-CreERT2 mice were mated to Eln(+/-) mice also carrying the multi-color ROSA26R(Rb/Rb) reporter. As described in step 1, plugs were checked, pregnant dams were induced with a single tamoxifen injection (1.5 mg) at E12.5, and they were sacrificed at E18.5. Embryos were harvested, fixed, frozen, and genotyped. Transverse descending aortic cryosections were cut. Sections from Eln(+/-), Acta2-CreERT2, ROSA26R(Rb/+) embryos indicate that the excess inner layer SMCs that accumulate in Eln-null mice (starting after E15.5) are marked by multiple colors (Figure 1) and hence derive from multiple alpha-smooth muscle actin (SMA)+ cells that are present at ~E12.5. In the E12.5 aorta, SMA+ cells are limited to the tunica media.
To generate Eln(-/-) embryonic aortas for explants, male and female Eln(+/-) mice were mated. Using methods described in step 2, plugs were checked and pregnant Eln(+/-) dams were sacrificed at E15.5. The aortas were isolated from Eln(-/-) embryos and cultured for 0 or 18 h in the presence of an anti-integrin β3 blocking antibody or an IgG1 control. Aortas were then fixed, frozen, and cryosectioned, and cryosections were stained for SMA (SMC marker), CD31 (endothelial cell marker), and nuclei (DAPI) (Figure 2). The results demonstrate that within 18 h of culturing, the E15.5 Eln(-/-) aorta becomes hypermuscular and stenotic. This process is attenuated by an anti-integrin β3 blocking antibody.
SMCs were isolated from the aorta of wildtype mice at P0.5. As described in step 3, SMCs were isolated from the dissected neonatal aorta by enzyme digestion and were cultured. Cells were passaged, and passage 3 cells were stained for SMC markers (i.e., SMA, smooth muscle myosin heavy chain (SMMHC), or transgelin (also known as SM22α)), CD31, and nuclei (DAPI) (Figure 3). Most of the cultured cells express SMC markers but not CD31.
To evaluate whether pharmacological inhibition of integrin β3 can attenuate hypermuscularization and stenosis of the Eln(-/-) aorta in vivo, we continuously infused the inhibitor cilengitide because of its short half-life in plasma. Eln(+/−) males and females were mated, and, as described in step 4, osmotic mini-pumps loaded with cilengitide were implanted in pregnant dams at E13.5. At P0.5, pups were euthanized and genotyped, and their aortas were analyzed. Cilengitide treatment significantly attenuates aortic stenosis and hypermuscularization in Eln(-/-) mice (Figure 4) without altering the wildtype aorta9. Thus, anti-integrin β3 is a potentially promising noninvasive approach to treat elastin aortopathy.
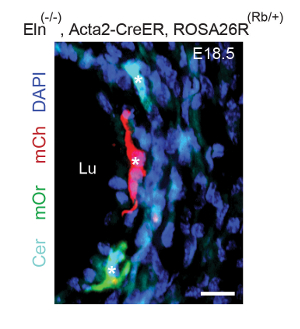
Figure 1. Excess SMCs in the Eln(-/-) Aorta Derive from Multiple Pre-existing SMCs. Dams pregnant with Eln(-/-), Acta2-CreERT2 embryos also carrying the multicolor Cre reporter ROSA26R(Rb/+) were induced with a single intraperitoneal injection of tamoxifen (1.5 mg) at E12.5. Dams were sacrificed at E18.5, and transverse sections of the descending aortas of the embryos were analyzed, with staining for DAPI and the direct fluorescence of Rb colors, Cerulean (Cer), mCherry (mCh), and mOrange (mOr). Excess SMCs accumulate in the inner aspect of the Eln-null aorta after E15.518. The excess inner-layer SMCs include cells of multiple colors (asterisks), indicating polyclonality. Lu, aortic lumen. Scale bar, 10 µm. Reprinted from Misra et al., 20169. Please click here to view a larger version of this figure.
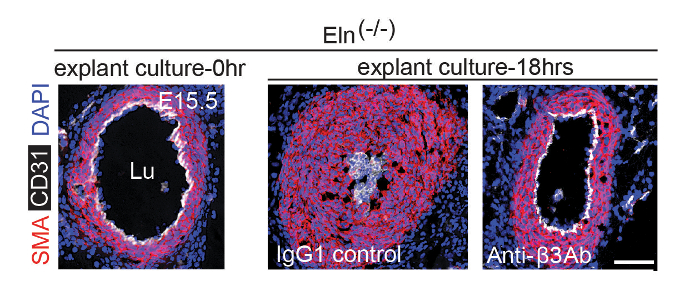
Figure 2. Anti-αvβ3 Integrin Blockade Attenuates Hypermuscularization and Stenosis of Eln(-/-) Aortic Explant. At E15.5, pregnant dams were euthanized and the aortas of Eln(-/-) embryos were harvested. Isolated aortas were either fixed immediately or cultured in the presence of an isotype control IgG1 or an integrin αvβ3 blocking antibody for 18 h prior to fixation. Fixed aortas were stained for CD31, SMA, and nuclei (DAPI). Lu, aortic lumen. Scale bar, 100 µm. Reprinted from Misra et al., 20169. Please click here to view a larger version of this figure.
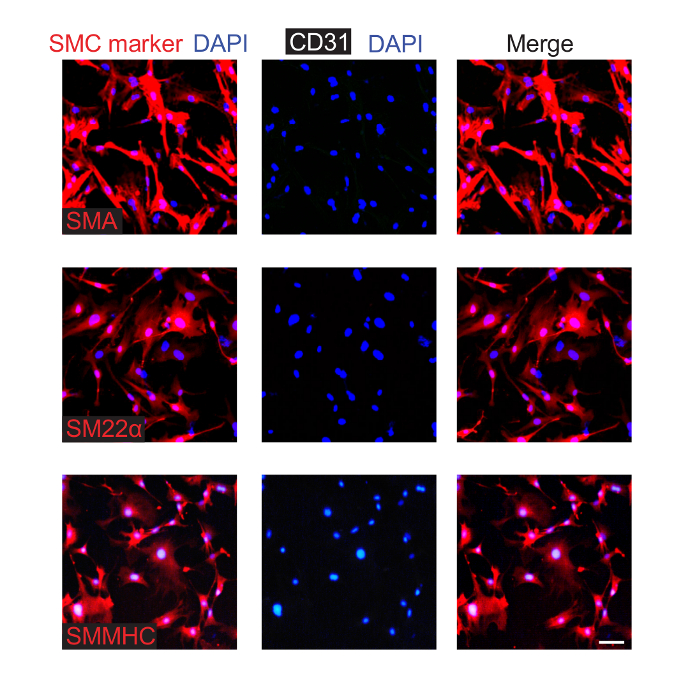
Figure 3. Isolated and Cultured Aortic SMCs from Neonatal Mice Express Smooth Muscle Markers. SMCs were isolated from P0.5 pups and were cultured. Cells from the third passage were fixed and stained for CD31 (EC marker), nuclei (DAPI), and an SMC marker (either SMA, SM22α, or SMMHC, as indicated). This analysis indicates that ~90% of the cultured cells express SMC markers, and none were observed to express CD31. Scale bar, 100 µm. Please click here to view a larger version of this figure.
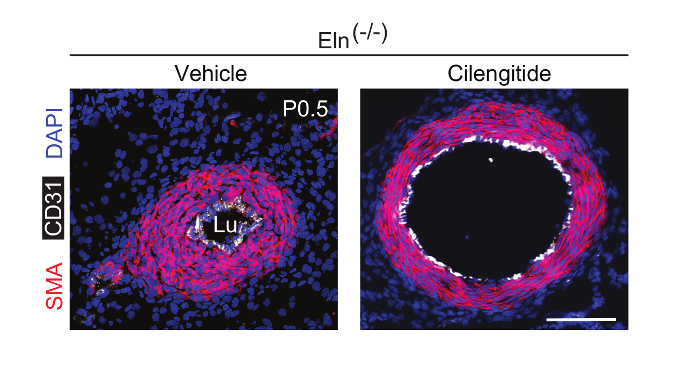
Figure 4. Continuous Infusion of Cilengitide In Vivo Attenuates Eln(-/-) Aortic Hypermuscularization and Stenosis. Male and female Eln(+/-) mice were crossed. The integrin β3 inhibitor cilengitide or vehicle (PBS) was continuously infused in pregnant dams using osmotic mini-pumps starting at E13.5. Transverse sections at P0.5 were stained for SMA (red), CD31 (white), and nuclei (DAPI, blue). Lu, aortic lumen. Scale bar, 100 µm. Please click here to view a larger version of this figure.
