A library of single point mutants in the SERTTC background was created to screen for thermostabilizing mutations. Individual mutants were generated using standard mutagenesis. The screening protocol utilizes transiently transfected HEK293S cells and a scintillation proximity-based thermostability screen to rapidly identity useful mutations for crystallization as outlined in Figure 1A. Plotting Tm values versus bound [3H]citalopram at RT reveals constructs with high thermostability and expression levels suitable for protein purification (Figure 1B). Three mutants (Y110A, I291A, and T439S) were combined to generate a highly stable construct (Figure 1C). Thermostability is also correlated with increased stability in short chain detergents necessary for the crystallization of the SERT-Fab complex.
The large scale expression of human SERT using baculovirus-transduced HEK293S GnTI– cells can take less than 2 weeks and can produce milligram quantities, as illustrated in Figure 2A. Use of the GFP-tagged SERTCC protein allows SERT to be conveniently followed during expression and purification by fluorescence (Figure 2B). Our purification strategy involved 1) solubilization of SERT bound to S-citalopram from HEK293S GnTI– cells in C12M in the presence of CHS as a stabilizing lipid; 2) binding of SERT to a Strep affinity matrix; 3) removal of contaminant proteins by extensive washing; and 4) elution of the functional SERT with buffer containing desthiobiotin (Figure 2C). The eluted protein is largely free of other detectable proteins by Coomassie blue staining and monodisperse as judged by FSEC (Figure 2D, E).
A similar strategy was taken to purify SERT with a Strep II tag which was used for reconstitution and immunization (Figure 3A, B). Incorporation of SERT into proteoliposomes boosts the serum half-life and stability of SERT and improves the likelihood of isolation of high-affinity antibodies. Furthermore, inclusion of lipid A, a component of the bacterial cell wall, serves as a potent adjuvant9. Multilamellar liposomes were prepared by the addition of buffer to a dried lipid mixture in glass tubes and resuspended in buffer. Extrusion of the liposomes through 200 nm pore size filters produces monodisperse unilamellar liposomal suspensions. The liposomes are then saturated with detergent followed by the addition of purified SERT in detergent. Finally, detergent is removed by addition of hydrophobic absorption resin to the lipid:detergent mixture. Additional ligand should be added to the reconstituted sample to select for antibodies that recognize the antidepressant bound conformation. The presence of SERT in the proteoliposomes should be confirmed by solubilizing a small sample with SDS-PAGE loading dye or C12M and running on SDS-PAGE and FSEC (Figure 3C, D).
Hybridoma cell lines expressing SERT antibodies can be screened for high-affinity binders which recognize 3D epitopes. These properties are crucial to the eventual success of crystallization, as the antibody must remain firmly bound to a structured region to promote crystal packing of homogeneous, well-ordered domains. In the first step, antibodies which recognize unstructured regions are identified. SERT is denatured and blotted onto a nitrocellulose membrane; antibodies which bind denatured SERT will be western-positive and likely recognize linear epitopes. In Figure 4A, we show 2 examples of antibodies which are western-positive and likely not useful to promote crystallogenesis. In Figure 4B, the remaining western-negative antibodies are incubated with 100 nM SERT-GFP and separated by FSEC. Antibodies which bind SERT will shift the GFP-positive peak to an earlier position. The SERT-antibody complexes may be diluted further in detergent to determine if they can bind with nanomolar affinity followed by analysis by FSEC. Addition of serotonin results in conformational changes in the transporter and thus the antibodies can be rescreened to determine if they can specifically recognize the SSRI-bound conformation. In Figure 4C, the antibodies are shown to bind SERT in the presence of serotonin, showing that the epitope(s) do not change from the SSRI to substrate bound state. Finally, in Figure 4D combinations of antibodies are tested for their ability to bind distinct epitopes, resulting in a further leftward shift. Here the 15B8 or 8A11 antibodies recognize an epitope which is different from 8B6.
The 8B6 antibody was chosen for further structural analysis based on preliminary crystal screening with papain treated Fab. The genes of the 8B6 Fab were cloned into an insect cell expression vector. Fab can be expressed and secreted from Sf9 cells growing in suspension. The 8B6 Fab can be purified from Sf9 cell supernatant by His-tag affinity (Figure 5A, B) and cation exchange chromatography (Figure 5C, D) resulting in protein which appears free of contaminants on SDS-PAGE gels. In Figure 5E, the recombinant 8B6 Fab is shown to bind SERT and is used in subsequent biochemical and biophysical experiments.
The affinity purified SERTCC is digested with thrombin and EndoH and mixed with 8B6 Fab to form a complex in the presence of S-citalopram. The transporter-antibody complex is then separated by SEC in C8M (Figure 6A) and the peak fractions contain both SERT and Fab as shown by SDS-PAGE (Figure 6B). Use of C8M is crucial for crystal formation probably because the short chain detergent allows better packing between molecules in the crystal lattice. FSEC is employed to determine which fractions should be pooled for crystallization (Figure 6C); fractions which are not monodisperse and/or contain large amounts of free SERT or Fab should not be combined.
Prism shaped SERT-antibody crystals can be grown in the presence of S-citalopram using this protocol by hanging drop vapor diffusion (Figure 7A). The resulting crystals diffract X-rays to a resolution of 3.15 Å10 (Figure 7B).
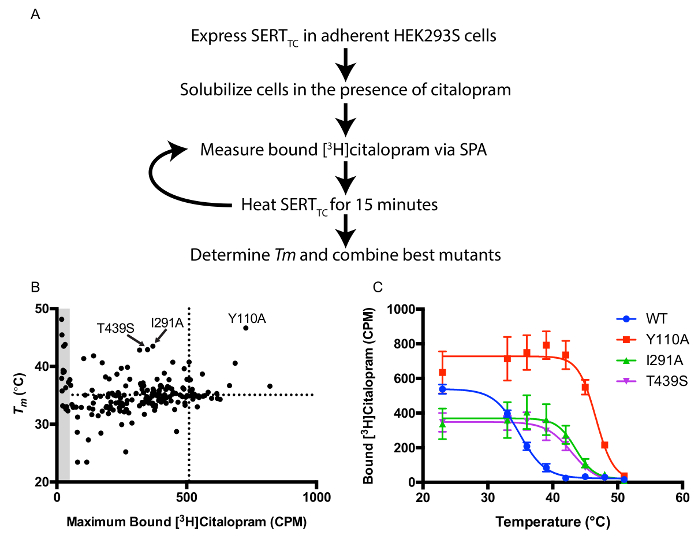
Figure 1: Scintillation Proximity-based Thermostability Assay. A. Overview of protocol for screening thermostability in the presence of [3H]citalopram. B. Maximum bound [3H]citalopram versus apparent melting temperature (Tm). Dotted lines represent values for the WT transporter. The 3 most thermostable mutants are labeled. Grey area represents mutants that have less than 10% of [3H]citalopram binding relative to WT and thus inaccurate Tm values due to a low signal-to-noise. C. Thermostability curves for WT SERTTC and the top 3 mutants. Error bars represent standard deviation (S.D.). Please click here to view a larger version of this figure.
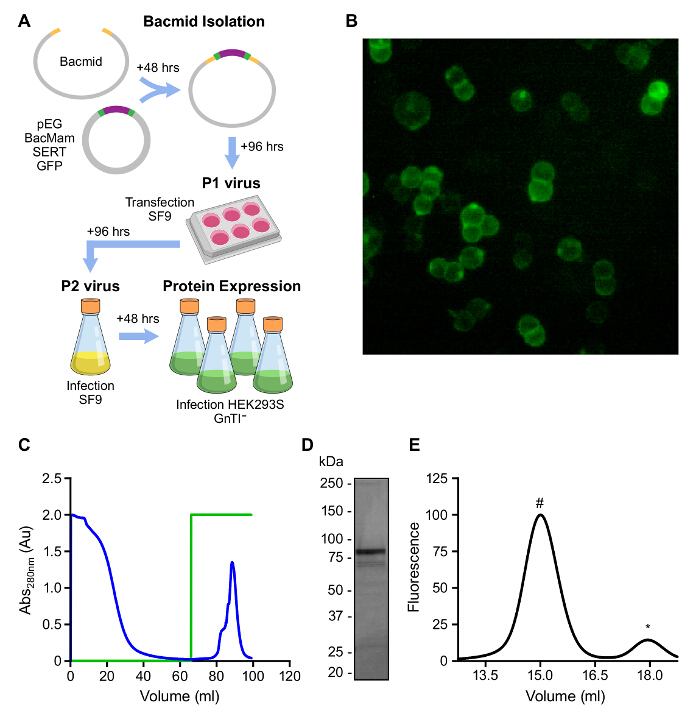
Figure 2: Overview of Mammalian Heterologous Protein Expression. A. Schematized overview of BacMam virus generation and expression of SERT in HEK293S GnTI– cells. B. HEK293S GnTI– cells expressing the SERTCC (GFP fluorescence). C. Elution profile of SERTCC on Strep affinity resin. Green trace represents the concentration of desthiobiotin, 0 – 100% (0 – 5 mM). D. Analysis of affinity purified SERTCC on a 4 – 15% SDS-PAGE gel. E. FSEC of affinity purified SERTCC detected by GFP fluorescence (Excitation: 480 nm; Emission: 510 nm). The peak eluting at 15 mL is SERT (#) and 18 mL is free GFP (*). Please click here to view a larger version of this figure.
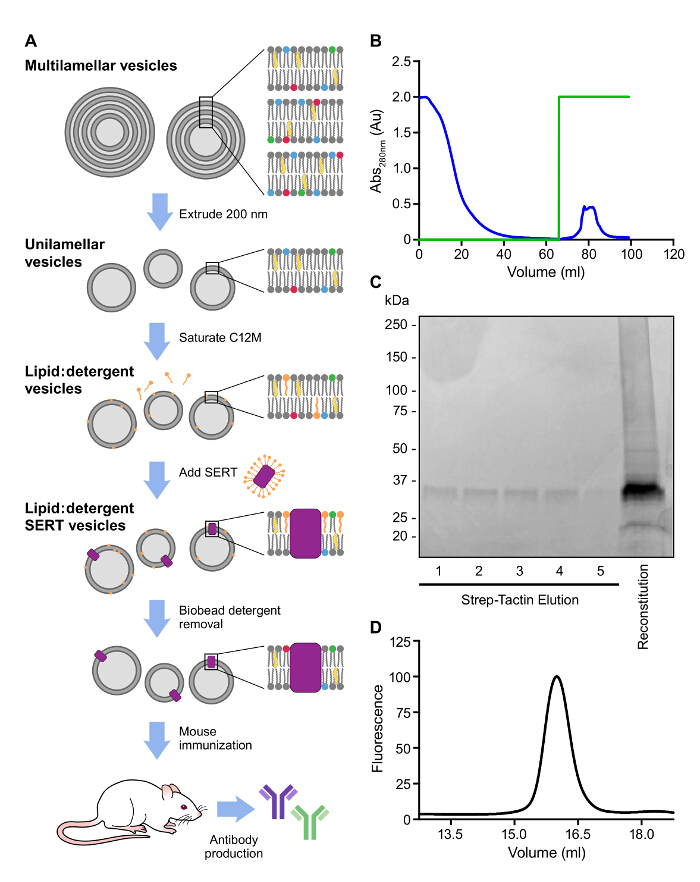
Figure 3: Representative Affinity Purification and Reconstitution of the SERTIC. A. Schematic overview of antibody generation. B. Elution profile observed at 280 nm of the affinity purification of SERTIC on Strep affinity resin. Green trace represents the concentration of desthiobiotin, 0 – 100% (0 – 5 mM). C. Analysis of affinity purified and reconstituted SERT on a 4 – 15% SDS-PAGE gel. D. FSEC of solubilized SERT following reconstitution. The fluorescence of tryptophan residues was used to detect SERT (Excitation: 280 nm; Emission: 335 nm). Please click here to view a larger version of this figure.
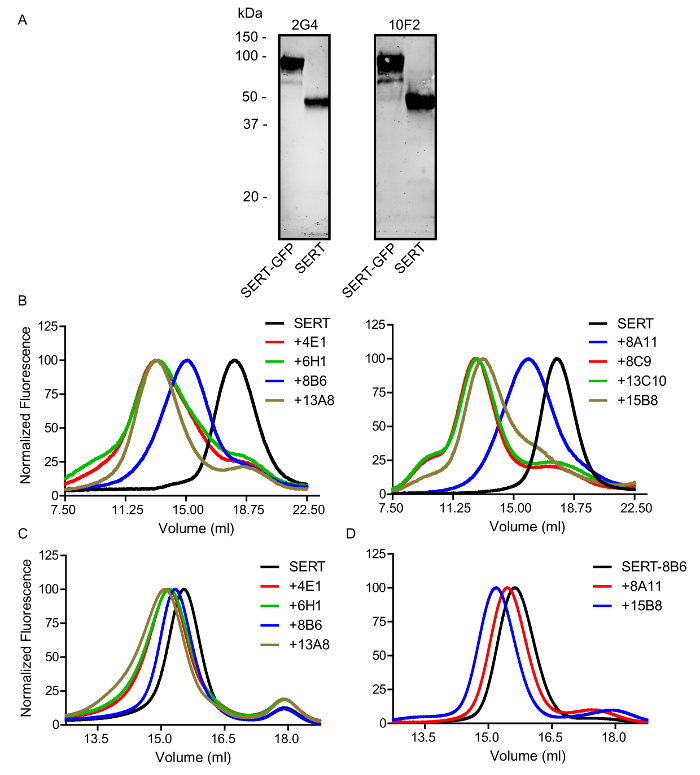
Figure 4: Analysis of Representative SERT Antibodies. A. Screening of antibodies by western blot. Approximately 1 µg of SERTCC with or without GFP was applied to a 4 – 15% SDS-PAGE gel and blotted onto a nitrocellulose membrane. Binding was detected using a goat anti-mouse antibody conjugated to IR Dye. 2G4 and 10F2 are western positive. B. Binding of antibodies to 100 nM GFP-tagged SERT and detection by FSEC detected using GFP fluorescence. C. Binding of selected Fabs to 100 nM GFP-tagged SERT in the presence of 1 mM serotonin. D. Binding of 8A11 or 15B8 Fabs to SERT-8B6 Fab. Minor peaks eluting at 18 mL are free GFP. Please click here to view a larger version of this figure.
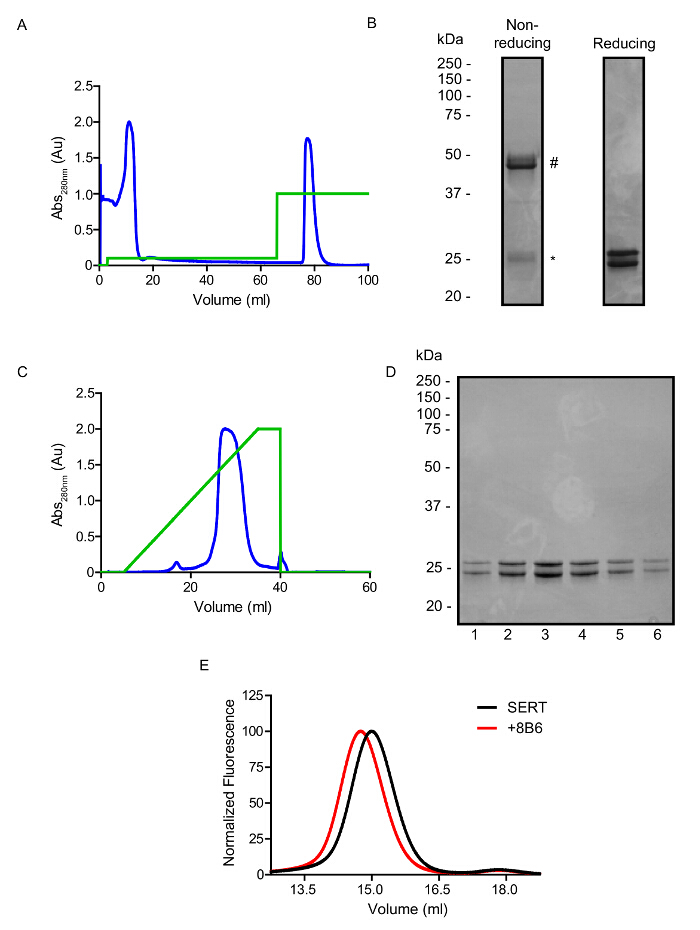
Figure 5: Representative Purification of the 8B6 Fab from Sf9 Cells. A. Elution profile observed at 280 nm of the purification of the 8B6 Fab by His-tag affinity chromatography. Green trace represents the concentration of imidazole, 0 – 50% (0 – 250 mM). B. Non-reducing and reducing SDS-PAGE gel after His-tag affinity purification. Protein which runs near 50 kDa is non-reduced Fab (#) and minor species at 25 kDa is reduced Fab (*). C. Elution profile observed at 280 nm of the purification of the 8B6 Fab by cation exchange displaying a single symmetrical peak which elutes under a linear sodium chloride gradient. Green trace represents the concentration of NaCl, 0 – 100% (0 – 500 mM). D. Analysis of the 8B6 Fab on a 12.5% SDS-PAGE gel following purification by cation exchange. E. Binding of the 8B6 Fab to 10 nM GFP-tagged SERT, detected using fluorescence of GFP. Please click here to view a larger version of this figure.
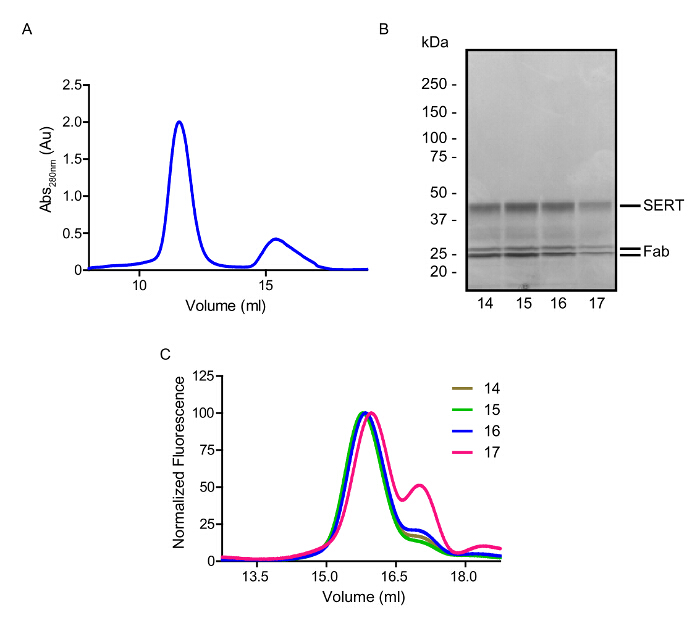
Figure 6: Representative Gel Filtration Chromatography of the SERT-8B6 Complex in the Presence of S-citalopram. A. Gel filtration elution profile of purified SERT-8B6 complex. Main peak eluting at 11.5 mL is the SERT-8B6 complex. Peak at 15 – 17 mL contains GFP and Fab. B. Analysis of the purified SERT-8B6 complex on a 4 – 15% SDS-PAGE gel. The positions of SERT and the heavy and light chains of the Fab are shown by a dash. C. FSEC of the size-separated fractions. SERT-8B6 complexes were detected using tryptophan fluorescence. Fraction 17 contains a larger amount of SERT which did not complex with Fab. Please click here to view a larger version of this figure.
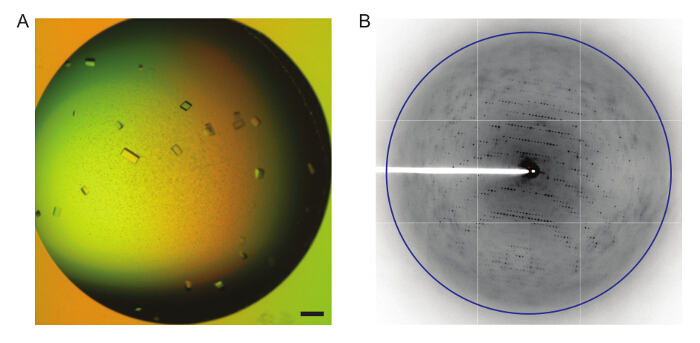
Figure 7: Crystallization of the SERT-8B6 Complex Bound to S-citalopram. A. Light microscopy of parallelepiped shaped crystals of the SERT-8B6 complex after 2 weeks of growth. Scale bar equals 200 µm. B. SERT-8B6 crystals diffract X-rays to 3.15 Å. Blue ring represents 3.15 Å. Please click here to view a larger version of this figure.
Table 1: A Crystallization Screen for the SERT-8B6 Complex Bound to S-citalopram. Please click here to download this table.
