Using AAVS1-specific ZNFs, fully characterized hESC/iPSC master cell lines containing heterotypic FRT target sequences were generated and described13. The FRT-containing master cell lines maintained pluripotency and genome integrity after the ZFN treatment and stably expressed GFP in vitro and in vivo. RMCE is performed by cotransfection of the master cell lines with FLPe recombinase and RMCE donor vectors (Figure 1A). RMCE vectors contain identical FRT sequences to the ones in the AAVS1 master cell line flanking the transgene. Figure 1B monitors RMCE using the constitutively expressing tdT pZ:F3-P CAGGS tdTPH-F. After transfection, hPSC form small groups of cells evenly distributed over the iDR4 MEF layer, and transfected hPSC express both GFP as well as transient tdT (Figure 1B, D 2). Cells are allowed to recover for 2 – 3 d post-transfection before starting selection. Positive selection is first started using a low dose of puromycin in order to favor RMCE donor insertion. Puromycin is applied gently at the start, because initial massive cell death could lead to single cells undergoing recombination, making them unable to survive the process. In the following days, puromycin concentration needs to be raised stepwise up to the optimal dose in order to allow the recombinant cells to grow into small colonies while the nonrecombined cells gradually die.
3 – 4 d after initiating positive selection (around D 6 post-transfection), negative selection is begun in order to select only for FRT-mediated recombination events. RMCE causes the loss of the thymidine kinase (Tk) suicide gene and thus the sensitivity to Fialuridine (FIAU). Negative selection applied at this point, in which small clusters of 2 – 4 GFP–/tdT+ (RMCE) cells are able to withstand optimal concentrations of FIAU (Figure 1B, D 6), prevents random integration, because in such events, the Tk gene in the AAVS1 locus remains unaffected. Occurrence of simultaneous FRT-mediated and random integration is highly unlikely, as demonstrated by the absence of random integration events later during the characterization (Figure 2B).
The mixed RMCE GFP–/tdT+ colonies continue to grow, while becoming more homogeneous, so that by D 9 – 10 post-transfection, some not fully selected mixed RMCE colonies can be found (Figure 1B). GFP–/tdT+ RMCE colonies are present only when the both RMCE donor and FLPe (2:1) are used, while RMCE does not occur in the absence of FLPe (2:0). Full selection with FIAU until day 13 – 15 post-transfection gives rise to a homogeneous GFP–/tdT+ culture. RMCE yields an average of 12.8 ± 6.8 (n = 6) PuroR/FIAUR colonies in 15 days13. Flow cytometry characterization of the newly generated RMCE line (the PuroR/FIAUR RMCE colonies combined in a nonclonal cell population) confirms that 100% of the cells present the RMCE GFP–/tdT+ phenotype (Figure 2A). When RMCE donors without constitutive expression of a fluorescent reporter are used, the resulting PuroR/FIAUR RMCE hPSC line is homogeneously GFP–.
PCR characterization of the nonclonal RMCE line demonstrates the full cassette exchange (no traces of the cassette of the master cell line can be detected) and that the selection program used generates random integration-free lines (both of the RMCE donor and the FLPe-expressing vector) (Figure 2B). These results were further proven by southern blot and, in addition, we demonstrated that the RMCE lines remain pluripotent13. This makes it no longer necessary to carry out either the genome-wide evaluation of random integration or the pluripotency test during routine RMCE experiments. In summary, using this protocol, hPSC transgenic cell lines in the AAVS1 locus free of random integration can be readily generated in 15 d with 100% efficiency and without the need of extensive characterization.
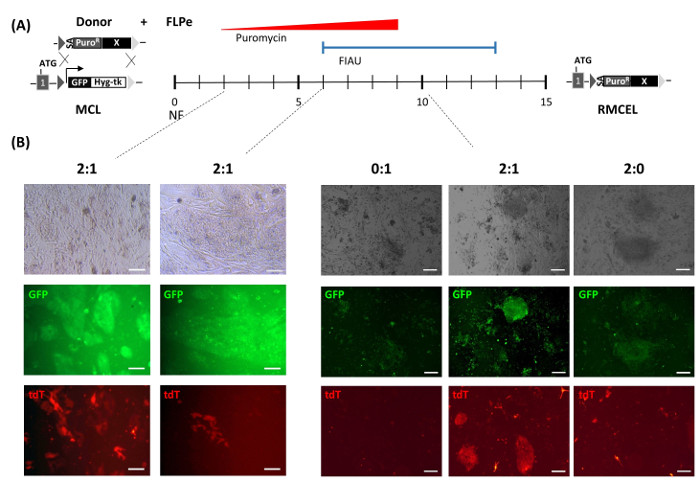
Figure 1: Schematic Overview of RMCE. (A) Timeline of the RMCE selection program. Left: the master cell line (MCL), harboring a FRT-flanked cassette (triangles) that express GFP and Hygromycin-tk (linked by 2A self-cleaving peptides), is transfected by nucleofection (NF) with the FLPe-expressing vector and the RMCE donor vector, which contains a promoterless Puromycin resistance gene (splicing acceptor – SA- PuroR) and a variable experimental cassette (X). Right: new RMCE line (RMCEL) obtained after completion of selection with Puromycin (red triangle) and FIAU (blue line). (B) RMCE using a constitutive tdT-expressing donor and different ratios of donor DNA and FLPe DNA (0:1, 2:1, 2:0) monitored by fluorescence microscopy (GFP – green fluorescence, tdT – red fluorescence) at different time points. D 2 and 6: scale bars represent 100 µm. D 10: scale bars represent 200 µm. Please click here to view a larger version of this figure.
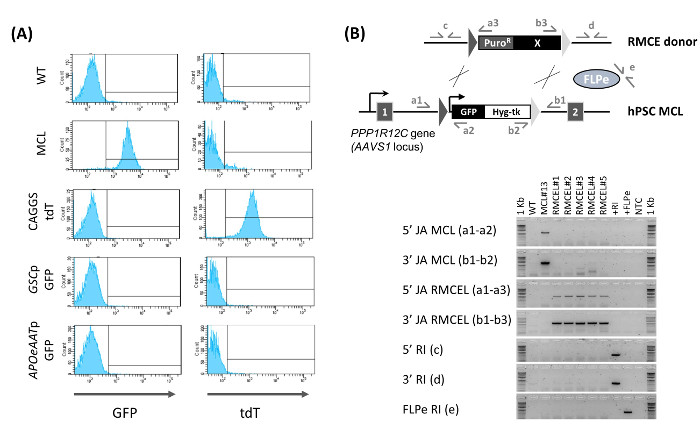
Figure 2: Characterization of RMCE Lines. (A) Determination of RMCE by flow cytometry. GFP and tdT expression is shown for the WT line, master cell line (MCL), and newly generated RMCE line at D 15 post-nucleofection using a constitutive tdT-expression vector (CAGGS tdT), or donors with GFP driven by a GOOSECOID promoter (GSCp GFP, definitive endoderm marker) or an AAT promoter (APOeAATp GFP, hepatocyte marker). (B) Confirmation of RMCE (5'/3' JA RMCEL), absence of the master cell line's (MCL) cassette (5'/3' JA MCL), and lack of random integration (5'/3'/FLPe RI) by PCR using the depicted primer pairs. DNA from WT, MCL, RMCE lines (1 – 5), donor RMCE vector (+RI), FLPe expressing vector, and no template control (NTC) were used. Figure modified from (Ordovas et al., 2015)13. Please click here to view a larger version of this figure.
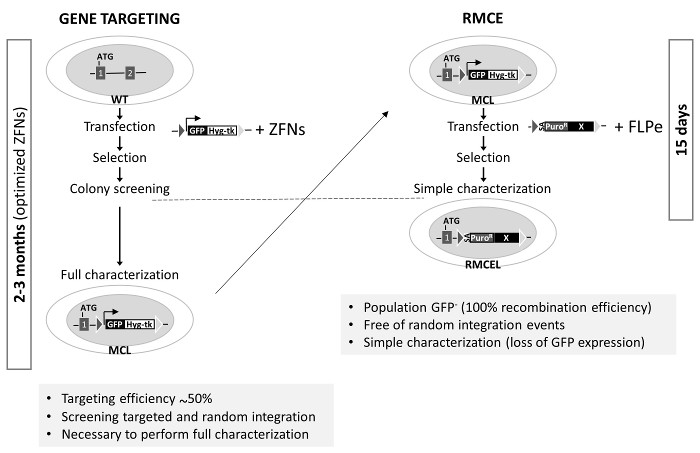
Figure 3. Generation of the RMCE-suitable Master Cell Line and Comparison of Gene Targeting with RMCE in AAVS1. Left: Gene targeting uses unmodified WT cells that are cotransfected with a donor (cassette described in Figure 1A for the generation of the master cell line, MCL) and specific optimized ZFNs. The process to complete the full characterization takes up to 3 months. Right: RMCE is performed by cotransfection of the donor with FLPe vectors, and a fully characterized line is generated in 15 d. Gene targeting efficiency in AAVS1 is from previous studies1,2. Please click here to view a larger version of this figure.
| Media | Components | Final concentration |
| hESC medium | DMEM-F12, HEPES | 80% |
| Serum replacement medium | 20% | |
| L-Glutamine | 146 mg/mL | |
| MEM Non-Essential Amino Acids Solution (100x) | 1x | |
| 2-Mercaptoethanol | 0.1 mM | |
| Penicillin/Streptomycin | 0.1% | |
| Human basic FGF | 4 ng/mL | |
| iMEF culture medium | DMEM High Glucose | |
| Fetal Bovine serum (FBS) | 15% | |
| Penicillin-Streptomycin | 0.1% | |
| L-Glutamine 200 mM | 4 mM | |
| MEM Non-Essential Amino Acids Solution (100x) | 2x | |
| 2-Mercaptoethanol | 0.1 mM | |
| Plating medium | hESC medium | 100% |
| ROCK inhibitor (Y-27632) | 10 µM |
Table 1. Media composition.
| Assay | Forward | Reverse | Amplicon | PCR Cycle | ||
| 5’JA MCL | CACTTTGAGCTCTACTGGCTTC | CGTTACTATGGGAACATACGTCA | 1.1 Kb | 95 ºC, 5’ – [95 ºC, 30’’ – 68 ºC (-0.5 ºC/cycle), 1’ 30’’]X15 – [95 ºC, 30’’ – 58 ºC, 30’’ – 72 ºC, 1’ 30’’]X25 – 72 ºC, 5’ | ||
| 3’JA MCL | TAACTGAAACACGGAAGGAG | AAGGCAGCCTGGTAGACA | 1.4 Kb | 95 ºC, 5’ – [95 ºC, 30’’ – 68 ºC (-0.5 ºC/cycle), 1’ 30’’]X15 – [95 ºC, 30’’ – 58 ºC, 30’’ – 72 ºC, 1’ 30’’]X25 – 72 ºC, 5’ | ||
| 5’JA RMCEL | CACTTTGAGCTCTACTGGCTTC | CATGTTAGAAGACTTCCTCTGC | 1.1 Kb | 95 ºC, 5’ – [95 ºC, 30’’ – 68 ºC (-0.5 ºC/cycle), 1’ 30’’]X15 – [95 ºC, 30’’ – 58 ºC, 30’’ – 72 ºC, 1’ 30’’]X25 – 72 ºC, 5’ | ||
| 3’JA RMCEL | TTCACTGCATTCTAGTTGTGG | AAGGCAGCCTGGTAGACA | 1.5 Kb | 95 ºC, 5’ – [95 ºC, 30’’ – 68 ºC (-0.5 ºC/cycle), 1’ 30’’]X15 – [95 ºC, 30’’ – 58 ºC, 30’’ – 72 ºC, 1’ 30’’]X25 – 72 ºC, 5’ | ||
| 5’RI DONOR | GTACTTTGGGGTTGTCCAG | TTGTAAAACGACGGCCAG | 0.5 Kb | 95 ºC, 5’ – [95 ºC, 30’’ – 60 ºC, 30’’ – 72 ºC, 30’’]X25 – 72 ºC, 5’ | ||
| 3’RI DONOR | CCTGAGTTCTAACTTTGGCTC | ACACAGGAAACAGCTATGAC | 0.5 Kb | 95 ºC, 5’ – [95 ºC, 30’’ – 60 ºC, 30’’ – 72 ºC, 30’’]X25 – 72 ºC, 5’ | ||
| RI FLPe | CCTAGCTACTTTCATCAATTGTG | GTATGCTTCCTTCAGCACTAC | 0.65 Kb | 95 ºC, 5’ – [95 ºC, 30’’ – 60 ºC, 30’’ – 72 ºC, 30’’]X25 – 72 ºC, 5’ | ||
Table 2. Primer Sets used for PCR Genotyping. Modified from Ordovas et al., 201513.
