In order for a protein to be considered as a legitimate deacetylation target for any enzyme with deacetylation activity, both in vitro and in vivo deacetylation assays need to be performed to establish the interplay between the deacetylase and its substrate. For the in vitro deacetylase assay, the purification of both the deacetylase and the acetylated protein substrate is required before the assay can be done. Here we use the cytoplasmic sirtuin SIRT2 as the specific deacetylase to be studied. SIRT2 is a deacetylase enzyme and the use of a mutant which lacks the deacetylase activity is highly recommended as a negative control for the deacetylation assay. Using the purification procedure described in 1.1, both SIRT2 and SIRT2H187Y can be successfully purified at a final concentration of 1 µg/µl. This can be confirmed after running the purified proteins on a gel followed by staining (Figure 1A, upper) as well as after western blotting using an anti-Flag antibody since both SIRT2 and SIRT2H187Y are tagged with a Flag peptide (Figure 2A, lower). The same purification procedure can be followed for the purification of the protein substrate with some modifications. The protein needs to be acetylated before it can be used for the deacetylation assay which means that it needs to be purified in cells overexpressing the specific acetyl transferase or a mixture of HATs able to acetylate the protein. Using the purification procedure described in 1.2, the acetylated protein can be purified (Figure 1B, upper). To confirm that the protein is indeed acetylated and can be used for the in vitro deacetylation assay, western blotting using an anti Ac-K antibody is necessary to show increased acetylated levels of the purified protein in cells overexpressing the acetyl-transferases (Figure 1B, lower).
After successful protein purification, the in vitro deacetylation assay can be performed. Sirtuins, including SIRT2, are class III histone lysine deacetylases which are distinct from other deacetylases as they require NAD+ for their enzymatic function. Consistent with this, no deacetylation can be detected by western blotting using an anti Ac-K antibody in the absence of NAD+, regardless of the presence of catalytically active SIRT2 (Figure 2, lane 2 vs 1). On the contrary, decreased acetylated levels of the acetylated protein when both SIRT2 and NAD+ are present in the reaction suggest that the protein can be considered as a deacetylation target for SIRT2. In order to verify these results, different negative controls can be used. For example no difference in the deacetylation levels of the protein can be detected when a deacetylation deficient SIRT2 (SIRT2H187Y) is used instead of the catalytically active wild-type SIRT2 (Figure 2, lane 5 vs 4). Similar effect can be seen when a well-established sirtuin inhibitor, NAM, is added to the reaction mixture, implying that the decrease in the acetylated levels of the protein is mediated by the deacetylase activity of SIRT2.
Aberrant deacetylation is involved in several cellular processes and age-related diseases. Thus, a crucial step in deepening our knowledge regarding the interplay between a deacetylase and its substrate is to establish this interconnection in cells where this phenomenon may have a physiological impact. Moreover, checking deacetylation in cell culture systems in vivo allows the study of the deacetylation event in a cell or tissue specific context which can be more easily connected to a specific phenotype or outcome. This excludes the possibility that the detected in vitro deacetylation activity is artificial due to the presence of both the deacetylase and its substrate in the tube at the same time, which may never happen under normal physiological conditions in cells. For the in vivo deacetylation assay, cells overexpressing both SIRT2 and SIRT2H187Y as well as the protein substrate can be used following the procedures described in 2.1 and 2.2. Cell extracts prepared from these cells can be confirmed to express SIRT2, SIRT2H187Y, and the protein substrate by western blotting using specific antibodies. In the assay described here, all exogenously expressed proteins are tagged for ease of the performed experiments (Figure 3, lower panel to detect HA-tagged SIRT2/SIRT2H187Y and middle panel to detect Flag-tagged protein substrate). To determine whether the target protein is a deacetylation substrate of SIRT2, immunoprecipitation is performed using an anti-Flag antibody to pull down the target protein followed by western blotting using an anti Ac-K antibody. Decrease in the acetylated levels of the protein in cells expressing SIRT2 (Figure 3 upper panel, lane 3 vs 2) and failure to affect acetylated levels in cells expressing the deacetylase deficient SIRT2 mutant (Figure 3 upper panel, lane 4 vs 3) suggest that the acetylated protein can be a deacetylation target of the specific deacetylase in vivo. This can be confirmed further when SIRT2-mediated deacetylation is inhibited after treating cells with NAM (Figure 3 upper panel, lane 5 vs 3) establishing the specific deacetylase-substrate axis in the studied cells.
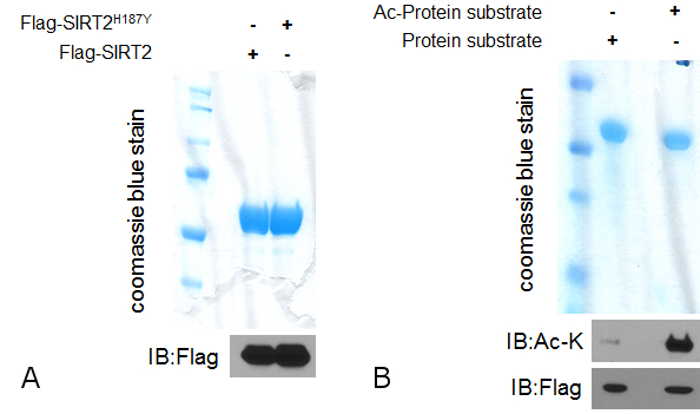
Figure 1: Purification of both the deacetylase and the acetylated protein-substrate to be used for the in vitro deacetylation assay. (A) 1 µg of both SIRT2 and SIRT2H187Y (deacetylation defective mutant) were run on a gel following the purification protocol as described in 1.1. The gel was stained with a commercially available staining solution and after destaining, both purified proteins can be detected (upper). Proteins can be detected after transferring in PVDF membrane and western blotting using an anti-Flag antibody (lower). (B) 1 µg of protein substrate and hyper acetylated protein substrate (alpha-enolase was used as a SIRT2 substrate based on mass spectrometry unpublished data generated in our lab) were run on a gel following the purification protocol as described in 1.2. The gel was stained with a commercially available staining solution and after destaining, the purified protein substrate can be detected (upper). Acetylation can be detected after transferring in PVDF membrane and western blotting using an anti-Ac-K antibody (middle). Total proteins can be detected using an anti-Flag antibody (lower). Please click here to view a larger version of this figure.
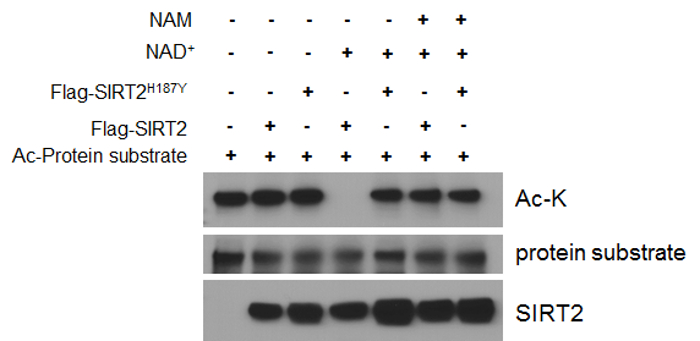
Figure 2: In vitro deacetylation assay. Purified acetylated protein substrate (alpha-enolase) is incubated with SIRT2 or SIRT2H187Y (deacetylation defective mutant) in the presence of NAD+ (lanes 4 and 5). Decreased acetylated levels are detected by western blotting using an anti Ac-K antibody in the presence of SIRT2 but not in the presence of SIRT2H187Y (lane 4 vs 5). No deacetylation activity is observed when NAD+ is not included in the reaction mixture (lane 2 vs 4) or when NAM is added to the reaction mixture (lane 6 vs 4). Please click here to view a larger version of this figure.
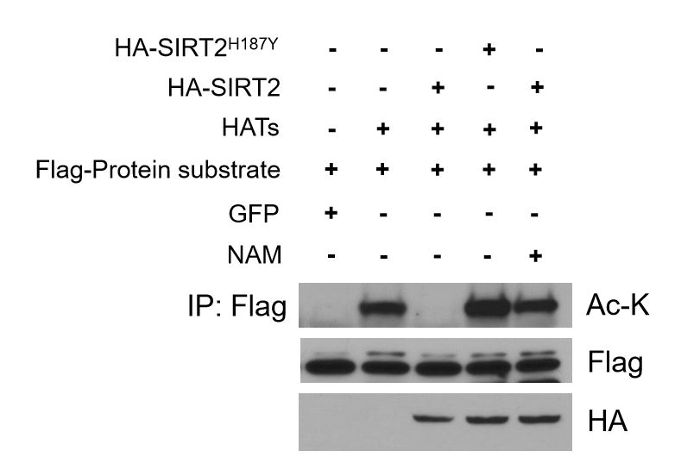
Figure 3: In vivo deacetylation assay. HEK293T cells stably overexpressing either SIRT2 or SIRT2H187Y were co-transfected with the protein substrate (alpha-enolase) and HATs to increase the acetylated levels of the protein (lane 2 vs 1). Deacetylation was checked in vivo after immunoprecipitation using an anti-Flag antibody to pull down the protein substrate and western blotting using an anti Ac-K antibody. Acetylation is significantly decreased in the SIRT2 overexpressing cells as compared to the SIRT2H187Y overexpressing cells (lane 3 vs 4). The SIRT2-mediated deacetylation of the protein substrate is inhibited when cells are treated with NAM (lane 5 vs 3). Please click here to view a larger version of this figure.
| Reactions | 1 | 2 | 3 | 4 (optional) | 5 (optional) |
| purified acetylated protein-substrate (10 μg) | + | + | + | + | + |
| buffer B | + | + | + | + | + |
| purified SIRT2 (2 μg) | – | + | + | – | + |
| NAD+ (1 mΜ) | – | – | + | + | + |
| purified SIRT2H187Y (2 μg) | – | – | – | + | – |
| NAM (10 mM) | – | – | – | – | + |
Table 1: Reaction ingredients.
| samples | overexpression | knockdown |
| 1 | pCDH-puro-GFP-empty vector | pLKO1 empty vector or si ctr |
| 2 | pCDH-puro-GFP-SIRT2-Flag | pLKO1 sh SIRT2 1 or si SIRT2 1 |
| 3 | pCDH-puro-GFP-SIRT2H187Y-Flag | pLKO1 sh SIRT2 2 or si SIRT2 2 |
Table 2: Protein samples.
