A typical experimental protocol, such as that used in this study, is illustrated in Figure 1. Following 20 consecutive days of VCD treatment, vaginal cytology was used to determine the presence or absence of cyclicity in the VCD-injected mice; vehicle-injected mice were also monitored. The stage in the estrus cycle was determined by the proportion of epithelial cells, cornified epithelial cells, and leukocytes in the vaginal smear (Figure 2A). The presence of only cornified epithelial cells indicated the mouse was in estrus, and therefore was still cycling. The estrous cycle in mice typically lasts 4 days, therefore VCD injected animals were considered acyclic after 15 days of persistent diestrus, in which vaginal cytology revealed a majority of leukocytes (Figure 2A). The estrous cycle of VCD-treated mice became irregular, and then ceased within 2-3 months after the start of VCD injections (Figure 2B). VCD ovarian tissue showed significant atrophy, compared to controls. At high magnification, we see a complete depletion of primary and primordial follicles (Figure 2C).
To test the effect of VCD-induced ovarian failure on exercise performance, we randomized 7 month old VCD- and vehicle-treated mice (two strains, C57BL/6 and B6C3F1) to either sedentary or cage wheel exercise groups 4-6 weeks following the cessation of cycling (Figure 1). Daily exercise values were recorded for time and distance for each exercised animal throughout the 4 week exercise period. Weekly averages of daily running values for distance, time and speed are shown in Figure 3 for C57BL/6 mice treated with vehicle or VCD. We did not see significant differences in exercise performance measured by average daily time, distance, and speed on the cage wheel between VCD- and vehicle-treated groups in either C57BL/6 or B6C3F1 mouse strain (Figure 4A). The calculated wheel-running speeds gradually increased over the 4-week running period as previously shown16, but were not significantly different between experimental groups. These studies indicate the impact of VCD-induced ovarian failure on voluntary cage wheel exercise transcends at least 2 mouse strains.
Following the duration of the voluntary cage wheel exercise protocol, body morphometrics were recorded and the hearts were rapidly excised and weighed. We have previously shown that voluntary cage wheel exercise induces an increase in cardiac mass16. Here, we also show cardiac hypertrophy in response to cage wheel exercise measured by absolute heart weight (HW) and HW normalized to tibial length (TL) (Table 1). The absolute heart mass and HW/TL ratio were significantly greater in vehicle- and VCD-treated mice when compared to sedentary counterparts. However, there were no measurable differences in cardiac hypertrophy between control and VCD-induced ovarian failure mice following voluntary cage wheel exercise.
Next, we used involuntary (forced) treadmill running to determine exercise capacity. We first subjected mice to a high-intensity protocol that increased average speed stepwise every 10 min until exhaustion. On average, the maximal speed of vehicle treated mice was 26.7± 0.95 m/min and the maximal speed of VCD treated mice was 28±1.4 m/min before exhaustion. We used lower-intensity running at 20 m/min (about 80% of maximal) to assay for endurance capacity. As shown in Figure 4B, no significant difference in high or low intensity exercise capacity was found between vehicle and VCD treated group, indicating that VCD-induced ovarian failure had no effect on mouse exercise capacity.
In conclusion, menopause induced by VCD treatment has no effect on either voluntary or forced running capacity, as well as the cardiac adaptive response to exercise.
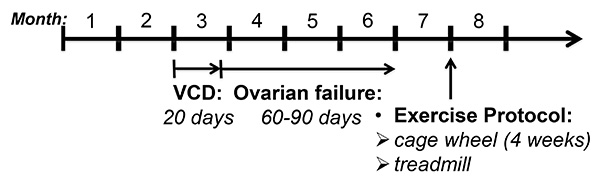
Figure 1. VCD-administration and experimental protocol. Mice were administered VCD (160 mg/kg) or sesame oil (vehicle control) starting at 2 months of age for 20 consecutive days. Following VCD or vehicle injections, estrous cycles were monitored and cyclicities were determined as described in the Methods section. Loss of ovarian function occurred within 60-90 days and was confirmed by vaginal cytology. At 7 months of age, mice were exposed to voluntary cage wheel exercise for 4 weeks or subjected to the treadmill exercise protocol.
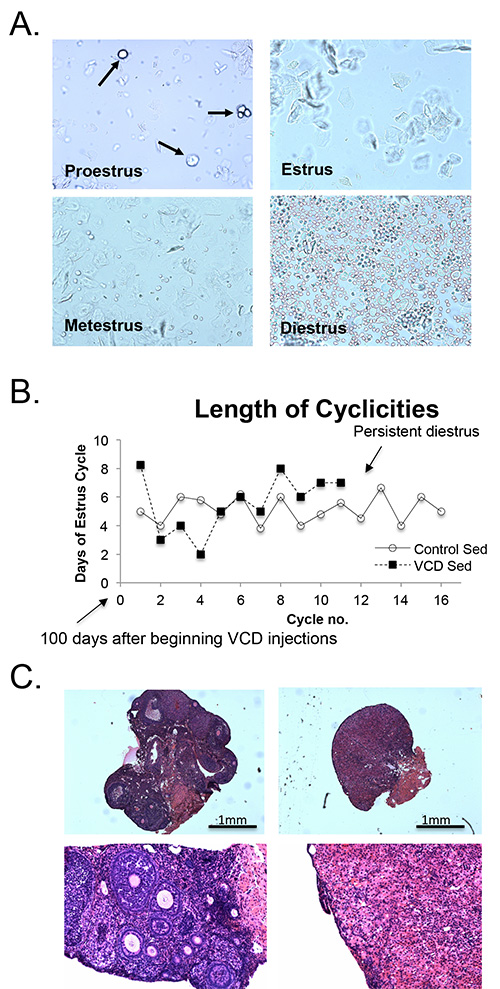
Figure 2. VCD-induced ovarian failure in mice. A: Cytology of an unstained vaginal smear from a vehicle injected mouse demonstrating the four stages of the estrous cycle: 1) proestrous, predominantly consisting of nucleated epithelial cells 2) Estrus, characterized by un-nucleated cornified cells; 3) metestrus, consisting of nucleated epithelial cells (arrows), un-nucleated cornified epithelial cells (arrows), and leukocytes; 4) diestrus, consisting primarily of leukocytes; and. B: Average length (days) of the estrous cycle of vehicle- and VCD-injected groups. One cycle is measured from the first day in estrus to the first day of the next period in estrus. Vaginal cytology began approximately 6 weeks following the onset of injections. The estrous cycles of VCD-injected animals become irregular, compared to vehicle-injected animals, and after 12 estrus cycles, cytology of all VCD injected animals no longer indicated cycling. C: H&E staining of ovarian tissue at 8 months of age. Cross-sections done at 5μm thickness for vehicle-treated (left panel) and VCD-treated (right panel) mice at 20X (top panel) and 200X (bottom panel) magnification. Ovarian tissue at 200X magnification shows a number of follicles in vehicle- compared to VCD-treated mice. Please click here to view a larger version of this figure.
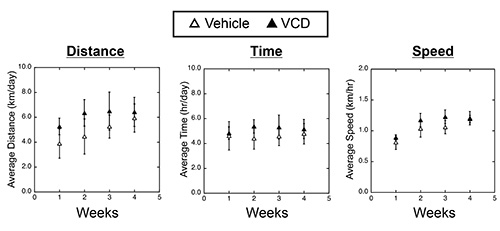
Figure 3. Voluntary cage wheel performance in vehicle- or VCD-treated C57Bl/6J female mice. Left Panel: Average running distance (km/day) for every 1 week period over the 4 week study period. Middle Panel: Average running time (hr/day) for every 1 week period over the 4 week study period. Right Panel: Average running speed (km/hr) for every 1 week period over the 4 week study period. Please click here to view a larger version of this figure.
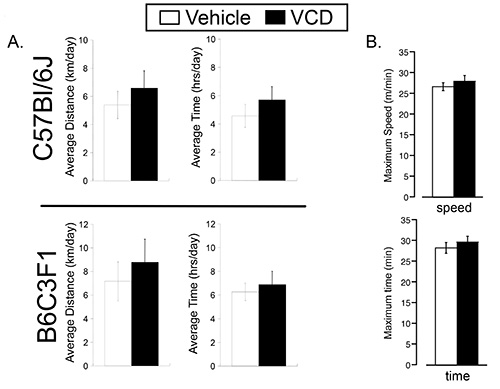
Figure 4. Voluntary cage wheel and treadmill performance in vehicle- and VCD-treated mice. A: (Top Panel) C57BL/6 average running distance (km/day) and time spent on the wheel (hrs/day) for every 24 hr over the 4 week study period. (Bottom Panel) B6C3F1 average running distance (km/day) and time spent on the wheel (hr/day) for every 24 hr over the 4 week study period. B: C57BL/6 treadmill running parameters. Maximum time (Top Panel) and maximum speed (Botoom Panel) using protocols detailed in the methods section.
