Flow cytometric assay for assessment of calcium mobilization in human PMNs
Incubating neutrophils with a concentration series of synthetic PSMα3 resulting in rapid activation as measured by calcium flux, which is shown by an increase in the signal in FL-1. Pre-incubation of synthetic PSMα3 with 0.01%, 0.1% or 1% human serum significantly inhibited the ability to elicit calcium fluxes (Figure 1).
Analysis of GFP expression in bacteria after phagocytosis by PMNs using fluorescence microscopy
Phagocytosed bacteria containing a PSMα-GFP reporter construct10 start to fluoresce green between 1 and 2 hr after phagocytosis, indicating expression from the PSMα promoter. Bacteria outside the neutrophils do not fluoresce, or show fluorescence only after the extracellular bacteria have formed dense microcolonies (Figure 2). These data indicate that the expression of PSMα is rapidly switched on when bacteria are phagocytosed by PMNs.
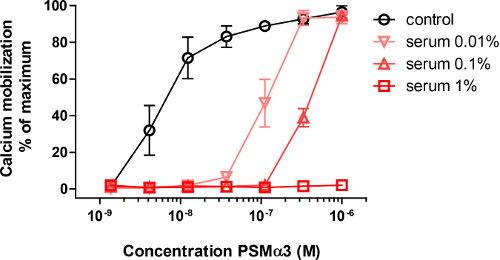
Figure 1. Neutrophil activation by PSMα3. Activation of Neutrophils by a concentration range of PSMα3, as measured by calcium mobilization. When very low amounts of serum are added the neutrophil activation is inhibited, and at 1% serum hardly any activation is visible at these PSMα3 concentrations (Adapted from reference 10).

Figure 2. Induction of expression of PSMα after phagocytosis. Neutrophils were allowed to phagocytose S. aureus containing a reporter construct of the promoter of PSMα fused to GFP. Approximately 1 hr after start of phagocytosis the intracellular bacteria start to fluoresce green, indicating expression of the psmα operon, whereas the bacteria outside the neutrophil (#) do not induce the psmα expression in this timeframe (Adapted from reference 10).
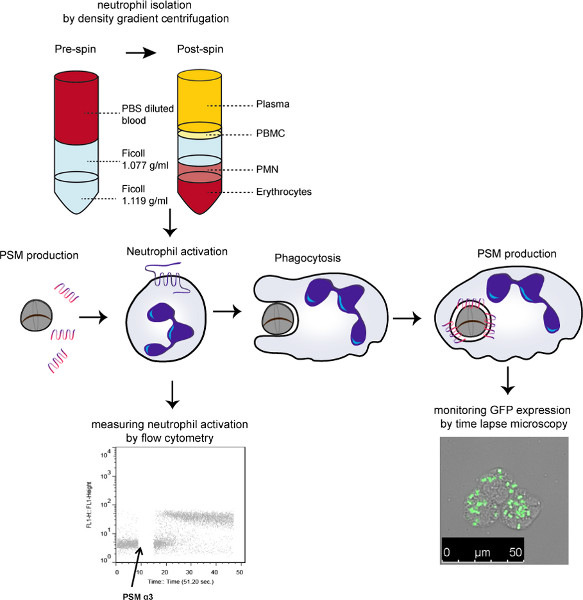
Figure 3. Schematic model of the experiments performed. Neutrophils were isolated and incubated with PSMs to measure the activation effect of these small amphipathic helices in a calcium mobilization assay. When serum was added, the PSMs were neutralized and no longer activated the neutrophils. To study the intracellular expression of the PSMs, a strain containing a fusion of the PSMα-promoter to GFP was mixed with serum and neutrophils to allow phagocytosis. GFP expression was followed using time-lapse fluorescence microscopy. Click here to view larger figure.
