1. Au NRs Preparation
Note: Au NRs can be synthesized by a number of recipes16, or purchased from commercial vendors.
- Measure the initial optical density (OD) of the Au NR solution via UV-Vis spectroscopy, by recording the absorption values from 300 nm to 1,000 nm with a resolution of 0.5-2 nm. Vary the volume of the solution to be used with the available cuvette.
- Evaluate the initial NP molar concentration with a suitable technique17 (e.g. UV-Vis spectroscopy, single particle inductively coupled plasma mass spectrometry, transmission electron microscopy) or use the concentration values provided by the vendor.
- Prepare a 5 ml stock solution by diluting the initial Au NR sample to reach an OD = 1. For the best repeatability, keep the OD constant for all of the tested samples. Follow the commercial vendor’s protocol for the composition of the diluting solvent. If unsure, use deionized water.
- Centrifuge 1 ml of the Au NR solution twice for 15 min at 7,800 x g to remove any chemical excess from the solution. Centrifugation cycles can vary in force and time (e.g. 20 min at 5,600 × g)18.
- Remove the supernatants and re-suspend the NRs in deionized water. As re-suspended particles might form aggregates in solution, prepare them daily to have the best results. Alternatively, store in a fridge for no longer than 1 week. Do not freeze.
- Before using under cell culture conditions, sonicate the Au NR solution for 5 min and then sterilize with UV light for 30 min (UV radiation intensity not less than 400 mW∙m-2 at 254 nm).
2. NG108-15 Neuronal Cell Line Culture and Differentiation
- For the cell culture medium, prepare 500 ml of sterile Dulbecco's modified Eagle medium (DMEM) containing 10% (w/v) fetal calf serum (FCS), 1% (w/v) L-glutamine, 1% (w/v) penicillin/streptomycin and 0.5% (w/v) amphotericin B.
Note: Supplements can be aliquoted, stored at -20 °C and added to the media on the day required. Cell culture medium can be refrigerated in a sterile condition for a maximum of 1 month. - For the cell differentiation medium, prepare 50 ml of sterile DMEM containing 1% (w/v) L-glutamine, 1% (w/v) penicillin/streptomycin and 0.5% (w/v) amphotericin B.
- Grow NG108-15 neuronal cells in 10 ml of cell culture medium in T75 flasks made of polystyrene in an incubator with humidified atmosphere (5% CO2 at 37 °C). Normally, seed 1.5-2 × 105 cells in each flask to be ready in 3-4 days. Change cell culture medium every two days.
Note: To prevent genetic drifts or variation, do not use cells older than passage 21 for experiments. - When 70-80% confluent in culture, change the medium with warm fresh cell culture medium. Mechanically detach the cells by gently knocking the bottom of the confluent flask. Do not use trypsin.
- Centrifuge the cell suspension for 5 min at 600 x g and re-suspend the cell pellet in 2 ml of warm cell differentiation medium.
- Seed 2 × 104 cells/cm2 in a tissue culture polystyrene 96 well plate with 200 µl of cell differentiation medium. Incubate the experiment for 1 day at 5% CO2/37 °C.
- Add between 3.2 × 109-4.2 ×1 010 particles/ml of Au NR solution on day 2 and incubate it for additional 24 hr. Do not add the particles for the control experiments.
Note: As an alternative control, Au NPs with a well-differentiated peak absorption wavelength can be used for comparison purposes14. For a more consistent cell behavior, keep the cell density constant and do not modify the well surface prior to cell seeding.
3. Neurite Outgrowth Enhancement
- Couple the laser with a single mode optical fiber (numerical aperture = 0.13) and terminate it with a fiber connector (FC connectors are convenient and commonly available). Measure the output laser power with a standard power meter. To obtain the most effective results, match the peak wavelength of the laser to the plasmon resonance peak of the NRs.
- On day 3 following NR incubation, fix the FC connector to the well. Irradiate samples and controls at RT for 1 min in continuous wave, for different laser powers. Allow the culture to proceed for 3 additional days at 5% CO2/37 °C. Repeat the laser irradiation for a minimum of 3 independent measurements.
Note: Different irradiation times and pulse frequencies may be selected, depending on the application. - Characterize the laser in terms of beam diameter and laser irradiance (W∙cm-2).
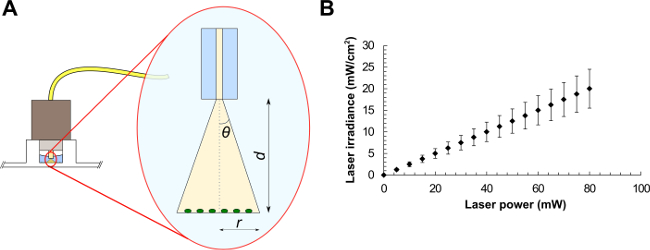
- For a standard single mode fiber, use NA = n∙sinθ, where n is the refractive index of the medium in use and θ is the half-angle of the cone of light exiting the fiber (see Figure 1A). From trigonometry, r = tanθ∙d, where r is the beam radius and d is the distance between the fiber and the sample. The FC connector matches the diameter of the well, therefore illuminating the sample at a distance d = 2.70 ± 0.20 mm. Light exits the fiber in water (n = 1.33), giving r = tan(sin-1(NA/n))∙d.
- Using the latter equation, calculate the laser beam radius, the corresponding beam area and the average laser irradiance (laser power divided by beam area) at the target (see the example graph in Figure 1B). These values represent the average irradiance over the illuminated area.
- Evaluate the errors for the measurements with the general theory of error propagation.
Note: The distance between the FC connector and the sample can be measured by photographs of the experimental arrangement and post-processing of the image using appropriate software (e.g. ImageJ).
- At day 5, remove cell differentiation medium from the experiments and fix the samples with 3.7% (v/v) formaldehyde solution for 10 min, then permeabilize the cells with 0.1% (v/v) Triton X-100 for 20 min.
- Add 3% (w/v) bovine serum albumin (BSA) to the samples for 60 min to block the unreacted protein binding sites. Label the samples for anti-βIII-tubulin overnight (5 µg/ml in PBS supplemented with 1% of BSA) at 4 °C.
- Incubate the cells for 90 min in the dark with an appropriate secondary antibody (e.g. TRITC-conjugated anti-mouse IgG antibody) using a concentration of 0.4-2 µg/ml in 1% BSA in PBS. Label the cell nuclei with DAPI (0.1 µg/ml in de-ionized water) for 10 min.
Note: Antibody conjugation and concentration might vary according to the company protocol. Wash samples with PBS twice for 5 min after each staining stage.
- Image samples by epifluorescence or confocal microscopy using at least a 20× objective. Choose the microscope filters accordingly to the secondary antibody. Select a DAPI filter (λEX = 358 nm; λEM = 488 nm) to visualize the cell nuclei.
- Analyze the pictures by assessing: i) the maximum neurite length (record the length from the tip of the neurite to the beginning of the cell body), ii) the number of neurites per neuronal cell (sum up all of the neurites per cell) and iii) the percentage of cells with neurites (divide the total number of cells expressing βIII-tubulin by the total number of cells with a positively stained nucleus)19.
4. Laser-induced Intracellular Ca2+ Imaging
- Prepare a 20% (w/v) stock solution of Pluronic F-127 by dissolving 2 g of solute in 10 ml of anhydrous dimethyl sulfoxide (DMSO). Heat the solution of Pluronic F-127 at 40 °C for about 20 min to increase the solubility. Prepare it in advance if stored at RT.
- On day 3 following NR incubation, prepare a balanced salt solution (BSS; 135 mM NaCl, 4.5 mM KCl, 1.5 mM CaCl2 , 0.5 mM MgCl2 , 5.6 mM glucose, and 10 mM HEPES, pH 7.4) and supplemented it with 5 µM of Fluo-4 AM in DMSO and 0.1% (w/v) of Pluronic F-127 solution.
Note: BSS solution can be prepared in advance and refrigerated for a maximum of 1 month. Add the supplements (Fluo-4 AM and Pluronic F-127) on the day of the experiment. - Remove cell differentiation medium and replace it with the supplemented BSS solution. To obtain the best results with an inverted confocal microscope, incubate the cells in a micro-slide well.
- Load NG108-15 cells with Fluo-4 AM for 30 min in the dark at 5% CO2/37 °C. Following the incubation time, wash samples twice with BSS to remove any extracellular unloaded fluorophore.
- Couple the laser with a single mode optical fiber and cleave the tip using standard techniques. Observe the resulting tip under an optical microscope, to ensure a high-quality surface (i.e. the tip should be perpendicular to the fiber axis and flat upon microscopy inspection).
- Measure the output laser power with a standard power meter. Insert the light delivery fiber into an optical fiber holder and affix it to a micropositioner. Refer to Brown et al.20 for more details on optical fiber preparation and positioning.
Caution: Follow the general rules for laser safety during the measurement of the laser power (e.g. do not look directly into laser beam, wear laser safety goggles during handling, prevent stray light exposure to other lab users).
- Measure the output laser power with a standard power meter. Insert the light delivery fiber into an optical fiber holder and affix it to a micropositioner. Refer to Brown et al.20 for more details on optical fiber preparation and positioning.
- Connect an oscilloscope to the portable laser to monitor the optical modulation. Use a binary signal with variable frequencies (0.5-2 Hz) and pulse lengths (20-100 msec). Connect the laser using the modulation signal as transistor-transistor logic (TTL) input for the microscope, following the setup shown in Paviolo et al.13.
- Place the cells under an inverted confocal microscope and position the light delivery fiber 250 ± 50 µm away from the target cell in transmission illumination mode. At this distance, use the equations presented in 3.3.1 to calculate the beam radius at the target.
- Use an argon-ion laser (488 nm) to excite the internalized Fluo-4 AM dye (λEX = 494 nm; λEM = 516 nm) and the synchronized LD for the excitation of the endocytosed NRs. Perform the imaging at RT of samples and controls using at least a 40× objective by collecting time-series scans with 256 × 256 pixels/frame resolution in roundtrip mode with a frequency of at least 4 Hz. Perform a recording without any LD excitation to identify any argon-ion laser interference (baseline noise).
- Remove the background from the data using adaptively weighted penalized least-squares algorithms21. Plot the induced Ca2+ variations as a function of time. Analyze the transient peaks in the fluorescence intensity using image post-processing software by thresholding at a level three times the standard deviation of the baseline noise (3σ).
By using Protocols 1, 2, and 3 described here, a stimulatory effect on differentiation was observed in NG108-15 neuronal cells cultured with Au NPs (Au NRs, poly(styrenesulfonate)-coated Au NRs and silica-coated Au NRs) after laser exposures between 1.25 and 7.5 W·cm−2 . Confocal images of rhodamineB-labelled Au NRs demonstrated that the particles were internalized from day 1 of incubation12. The localization was predominantly observed in the cell cytoplasm, indicating that the preferred mechanism of uptake was via the cell body membrane12. The main morphological changes detected after inducing differentiation in NG108-15 neuronal cells were the arrest of proliferation, the expression of βIII-tubulin protein and the outgrowth of neurites, which were analyzed in terms of maximum length and number22.
Samples cultured with NRs showed a neurite length increase at the laser irradiance levels measured here (between 1.25 and 7.5 W·cm−2). Control samples (cells cultured without NPs and irradiated with the same laser power) showed no significant change in length. Using an irradiation dose of 7.5 W·cm−2, the final neurite length of NG108-15 cultured with Au NRs was roughly 36% higher (p <0.01) than the non-laser-irradiated samples. This behavior was not specifically linked to the surface chemistry of the NRs. These values were almost 20% greater than the neurites developed by NG108 -15 alone and exposed to the same value of laser exposure (p <0.05)12. These results are in line with previously published studies on PC12 neuronal cells cultured with piezoelectric nanotubes and irradiated with ultrasound radiation7.
Control experiments without Au NRs also showed some stimulatory effects of the 780 nm light in terms of percentage of neurons with neurites and number of neurites per neuron. This stimulation was more effective at lower laser powers (1.25 W∙cm-2) with a subsequent decrease at the highest laser energy (7.5 W∙cm-2)12. A moderate stimulation caused by the NPs without any laser irradiation (poly(styrenesulfonate)-coated and silica-coated only) was detected in the percentage of neurons with neurites12. These results are in line with recently published observations that gold nanoparticles can increase neuronal activity in vitro23,24. Figure 2 shows an example of epifluorescence images of differentiated NG108-15 neuronal cells cultured alone (Figure 2A) or with Au NRs (Figure 2B) and irradiated with different laser powers (indicated in the figure).
The potential for photo-generated intracellular Ca2+ release was assessed using pulsed NIR light in accordance with Protocols 1, 2, and 4. Calcium ions play an important role in different cellular activities, such as mitosis, muscle contraction and neurite extension25,26. In response to a stimulus, Ca2+ increases, oscillates and decreases, leading to the activation, modulation or termination of a specific cell function. Recently, Ca2+ transients have also been observed as a consequence of IR laser exposure in cardiomyocytes. In that work, the Ca2+ responses evoked after IR exposure exhibited lower amplitudes and faster recovery times than the spontaneous Ca2+ transients2. Figure 3 shows an example of NG108-15 neuronal cells loaded with Fluo-4 AM and imaged with a confocal microscope. Fluo-4 AM was observed to enter the cell membrane in a non-disruptive manner, resulting in a generally uniform distribution of the indicator across the cell cytoplasm. Only minor nuclear or cytoplasmic organelle staining was detected. As previously indicated, NRs were expected to be located intracellularly12. NG108-15 neuronal cells alone or cultured with Au NRs and poly(styrenesulfonate)-coated-Au NRs were exposed thereafter to laser irradiances between 0.07 J∙cm-2 and 370 J∙cm-2, with the laser frequency modulated in the range of 0.5-2 Hz.
Figure 4 shows representative examples of how the amplitude of the responses was mapped as a function of time. The amplitude of the Ca2+ response varied with the radiant exposure (Figure 4A–C) and was observed not to be consistently triggered by the laser pulses (Figure 4C)13. The most likely explanations were the transient depletion of intracellular Ca2+ stores attributed to incomplete Ca2+ loading2 or the different efficiency of NR internalization in the neuronal cells. When NRs were not used in culture (control experiments, Figure 4D), a stimulatory effect of the 780 nm light was also observed. However, this produced lower fluorescence amplitude peaks and lower probability of activation (detected in only 16% of the analyzed samples). Overall, a 48% probability of NR laser-induced cell activation was achieved and despite the background events due to the 780 nm light, exposure of NR-treated cells demonstrated higher stimulation efficiency with lower laser energy and higher peaks of response13. In fact, the calcium response was found to peak at 0.33 J∙cm-2 in the NR-treated cells. This was attributed to thermal inhibition13. During the experiments, no evidence of blebbing or any other form of cell membrane disruption was detected, which is consistent with the results of Huang et al. that reported cellular photodestruction with a relatively high power density of 19 W∙cm-2 applied for 4 min27. No spontaneous activity in the NR-treated cells was recorded without laser exposure.
Figure 1: Optical fiber experimental setup (A) and average laser irradiances as a function of the laser power for a laser beam of area equals to 0.4 mm2 (B). Beam parameters are (A): the half-angle of the maximum cone of light exiting the fiber (θ), the beam radius (r) and the distance between the optical fiber and the sample (d).
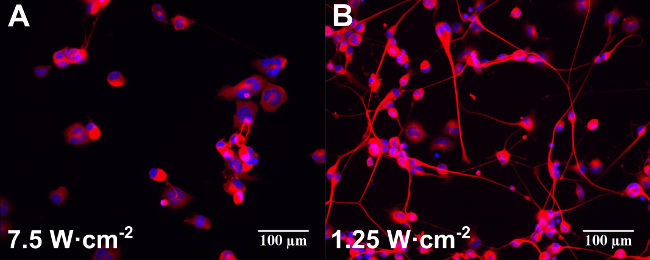
Figure 2: Examples of epifluorescence images of differentiated NG108-15 neuronal cells cultured alone (A) or with Au NRs (B) and irradiated with different laser powers (indicated in the figure). Samples were incubated for one day before laser irradiation. Cells were fixed and labeled for anti-β-III tubulin (red) and DAPI (blue) three days after laser irradiation. Scale bars are 100 µm.
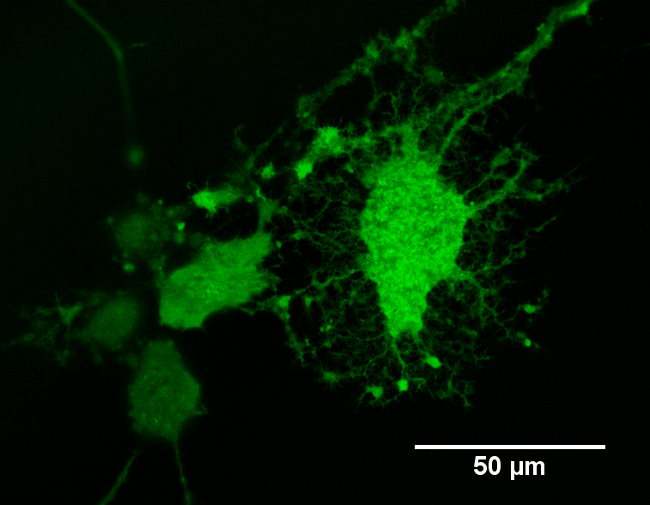
Figure 3: Example of differentiated NG108-15 neuronal cells loaded with Fluo4-AM Ca2+ indicator. The image was taken using an inverted confocal microscope with a 40X oil-immersion objective.
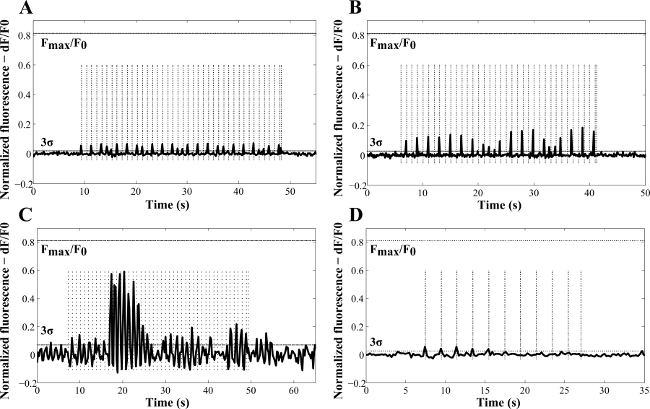
Figure 4: Representative examples of laser-induced Ca2+ variations as a function of time in NG108-15 neuronal cells cultured in serum-free conditions for three days with (A) poly(styrenesulfonate)-Au NRs, (B, C) Au NRs, and (D) without NRs (control sample). These results were obtained with laser pulses of 100 msec (A, C, D) and 50 msec (B). The frequencies used for the experiments (dashed vertical lines) were 1 Hz (A–C) and 0.5 Hz (D). The calculated radiant exposures were 69.4 J∙cm-2 (A), 34.7 J∙cm-2 (B), 0.37 J∙cm-2 (C) and 138.87 J∙cm-2 (D). Fmax /F0 indicates the maximum fluorescence increase detected in NG108 -15 neuronal cells, from calibration with ionomycin (reproduced with permission13).
