1. Preparation of Colon Tissue for GIMM
- To prepare a segment of distal colon for the Gastrointestinal Motility Monitor (GIMM), first place the isolated colon in ice-cold Krebs solution (121 mM NaCl, 5.9 mM KCl, 2.5 mM CaCl2, 1.2 mM MgCl2, 25 mM NaHCO3, 1.2 mM NaH2PO4, and 8 mM glucose; aerated with 95% O2/5%CO2). Clear away remaining mesentery from the outer wall and make a small incision in the oral end so it can be distinguished when placed into the organ bath. Note: tissue may remain in iced Krebs solution for up to 2 hours prior to experimentation.
- Next, position the inflow and outflow conduits in the organ bath so they are outside of the camera field to prevent interference with the image acquisition. Continuously perfuse the organ bath with prewarmed (37°C) oxygenated (95%, 5% CO2) Kreb’s solution at a flow rate of 10 ml/min.
- Keeping track of the oral vs. anal ends, pin a segment of distal colon (at least 5 cm) on either end in the organ bath, allowing a small degree of laxity so that the segment can move freely up to 1 cm in the middle. The oral end should be positioned towards the researcher for ease of placing the fecal pellet. Colonic segments should be pinned in the same manner by the same researcher for every experiment within a given set of experiments because the length and tension of the segment affects the rate of propulsive motility, with longitudinal stretch decreasing the rate of transit (Dickson et al., 2007).
- Allow the preparation to equilibrate for at least 30 min.
2. Setting up GIMM and Data Acquisition
- In the GIMM system, the colonic segment in the perfusion chamber is illuminated from beneath. A digital video camera interfaced with a computer is positioned above the chamber. Ensure that both the light illumination source and GIMM software are turned on.
- After setting up a new experiment in the GIMM software application, begin the first trial by inserting an epoxy-coated fecal pellet into the oral end of the colonic segment to initiate peristalsis. Click on the camera toggle switch on the computer to turn on the camera and click on the record button to start recording. The movement of the pellet in the anal direction is recorded by the video camera and the digital movies are stored on a PC for later analysis. When the pellet has reached the end of the colonic segment, click the record button to stop recording.
- To obtain a control value for the rate of propulsion, start with a colonic segment from a healthy animal and without applying drugs. Conduct 3-5 trials in a single preparation, with a recovery period of 5 min between each run.
- To determine the effects of certain conditions or drugs on colonic motility, perform 3-5 trials/preparation for each experimental condition. In addition, perform each experiment on at least five different colons from at least five different animals.
- In the analysis of the digital movies, the rate of fecal pellet propulsion is calculated as the time it takes for a pellet to traverse X cm of the colonic segment.
3. Construction of Spatiotemporal Maps
- The digital videos acquired from individual GIMM runs can be converted to spatiotemporal maps using the custom GIMM software.
- On the horizontal axis, changes in colonic diameter are plotted over time by converting the image of the colon in each video frame to a silhouette, and calculating and converting the diameter along the entire length into grey-scale. The end result is that the pellet and areas of relaxation appear black, while areas of contraction appear white.
- The distance traveled by the pellet through the colon segment is represented on the vertical axis.
4. Representative Spatiotemporal Maps
- Shown here are representative spatiotemporal maps showing pellet motility in colonic segments under various experimental conditions. The y (vertical) axis represents pellet position over time, while the x (horizontal) axis represents the distance that the pellet progressed through the colon segment.
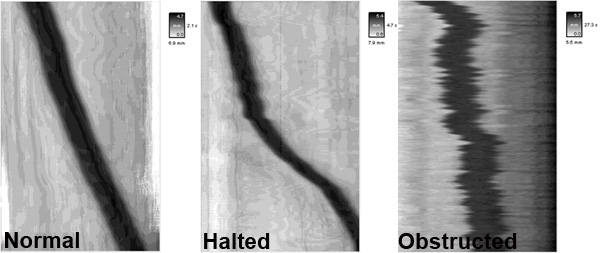
- In the spatiotemporal map from a control colon (untreated, normal), the pellet will progress linearly at a rate of ~2 mm/sec. In contrast, the administration of certain drugs or inflammation in the colon can result in disrupted motility patterns such as halted motility and obstructed motility.
- Results from GIMM can also be shown as graphs, in which the y-axis represents the distance in millimeters traveled by a fecal pellet and the x-axis represents time in seconds. For example, administration of DAMGO (D-Ala2, N-Me-Phe4, Gly-ol5), an opiate μ-receptor agonist, causes a decrease in propulsive motility in the colon.
