High-resolution x-ray microcomputed tomography (micro-CT) is an imaging method in which multiple radiographs (projections) of a sample are recorded from different viewing angles and later used to provide a virtual reconstruction of the sample1. This virtual object can then be analyzed, manipulated, and segmented, allowing non-destructive exploration in three dimensions2. Initially designed for medical analyses and later for industrial applications, micro-CT also offers the advantage of visualizing inner organs and tissues without the need for invasive procedures3. Like other forms of imaging, micro-CT works with a trade-off between the field of view and pixel size, which means that high-resolution imaging of large samples is nearly unattainable4. Advances in using high-energy X-ray sources (i.e., synchrotron) and secondary optical magnification are constantly being made, allowing the smallest resolution to reach under 100 nm5,6. Nevertheless, longer scanning times are necessary for large samples, increasing the chance of artifacts due to sample movement or deformation inside the scanner. Furthermore, micro-CT is generally limited by natural density variations within the sample and how the sample interacts with X-rays. While a higher X-ray dose is best for penetrating denser samples, it is less efficient in capturing variations in density within and between the sample and its surrounding medium7. On the other hand, a lower X-ray dose offers less penetration power and often requires longer scanning times but more sensitivity in density detection7.
These restrictions have long hampered the use of microtomography for plant sciences, given that most plant tissues are composed of light (non-dense) tissue with low X-ray absorption8. The first applications of micro-CT were focused on mapping root networks within the soil matrix9,10. Later, plant structures with more significant differences in tissue density, such as wood, began to be explored. This has allowed investigations of xylem functionality11,12, development of complex tissue organizations13,14, and interactions among plants15,16,17. The analysis of soft and homogeneous tissue is becoming widespread due to contrast agents, which are now standard procedure in preparations for micro-CT scanning of plant samples. However, protocols for contrast introduction can have different results depending on sample volume, structural properties, and the type of solution used8. Ideally, the contrast agent should enhance distinction among different tissues, enable tissue/organ functionality evaluation, and/or provide biochemical information about a tissue18. Therefore, adequate sample treatment, preparation, and mounting before scanning become crucial for any micro-CT analysis.
Micro-CT of the parasitic plant haustorium
Parasitic flowering plants represent a distinct functional group of angiosperms characterized by an organ known as haustorium19. This multicellular organ, a developmental hybrid between a modified stem and a root, acts on the host's attachment, penetration, and contact by a parasite20. For this reason, the haustorium is considered to "embody the very idea of parasitism among plants"21. A detailed understanding of this organ's development, structure, and functioning is crucial for parasitic plant ecology, evolution, and management studies. Nevertheless, parasitic plants' overall complexity and highly modified structure and haustoria often hinder detailed analysis and comparison. Haustorium connections are also usually extensive and not homogenous in tissue and cell distribution (Figure 1). In this context, while working with small tissue fragments allows easier manipulation and higher resolution, it can lead to erroneous conclusions about the three-dimensional architecture of complex structures, such as the parasitic plant haustorium.
Although there is a vast literature on haustorium anatomy and ultrastructure for most parasitic plant species, the three-dimensional organization and the spatial relationship between parasite and host tissues remains poorly explored17. In a recent work by Masumoto et al.22, over 300 serial semi-thin microtome sections were imaged and reconstructed into a three-dimensional virtual object representing the haustorium of two parasite species. This method's excellent level of detail provides unprecedented insights into the haustorium's cellular and anatomical 3-D structure. However, such a time-consuming technique would forbid a similar analysis in parasites with more extensive haustorium connections. The use of micro-CT emerges as an excellent tool for three-dimensional analysis of complex and often bulky haustoria of parasitic plants. Although not a substitute for detailed anatomical sectioning and other complementary forms of microscopy analyses17,23, results obtained via micro-CT scanning, especially for large samples, can also serve as a guide to direct the sub-sampling of smaller segments, which can then be analyzed using other tools, such as confocal and electron microscopy, or re-analyzed with high-resolution micro-CT systems.

Figure 1: Parasitic plants of different functional groups used in this protocol. Euphytoid parasite Pyrularia pubera (A), endoparasite Viscum minimum (B) with green fruits (dashed black circle), parasitic vine Cuscuta americana (C), mistletoe Struthanthus martianus (D), obligate root parasite Scybalium fungiforme (E). Segments of the host root (Hr) or stem (Hs) facilitate the application of contrast into the parasite haustorium (P). The presence of parasite mother root/stem (arrows) in the sample allows analysis of haustorium vessel organization. Rectangles indicate segments of the sample used for analysis. Scale bars = 2 cm. Please click here to view a larger version of this figure.
As micro-CT becomes an increasingly popular technique in plant sciences, there are guides, protocols, and literature dealing with sample scanning, three-dimensional reconstruction, segmentation, and analysis3,10,24. Thus, these steps will not be discussed here. As with any imagining technique, appropriate treatment and mounting of samples are a fundamental, albeit often being an overlooked procedure. For this reason, this protocol focuses on the preparation of haustorium samples for micro-CT scanning. More specifically, this protocol describes two approaches for introducing contrast agents into haustorium samples to improve visualization of different tissues and cell types in the haustorium, to facilitate the detection of parasitic tissue within the host root/stem, and to analyze parasite-host vascular connections in three dimensions. The preparations described here can also be adapted to the analysis of other plant structures.
Five species were used to better illustrate the convenience of the protocol described here. Each species represents one of the five functional groups of parasitic flowering plants, thus addressing specific points related to the functionality of each group. Pyrularia pubera (Santalaceae) was chosen to represent euphytoid parasites, which germinate in the ground and form multiple haustoria that connect the parasite to the roots of its hosts25. The haustoria created by these plants are often tenuous and easily torn apart from the host26 (Figure 1A), thus requiring a more delicate handling process. Endoparasites are represented here by Viscum minimum (Viscaceae). Species in this functional group are only visible outside the body of their hosts for short periods (Figure 1B) and live most of their life cycles as significantly reduced and mycelial-like strands of cells embedded within host tissues25. A third functional group comprises parasitic vines, which germinate on the ground but form only rudimentary roots, relying on multiple haustoria that attach to the stems of host plants25 (Figure 1C). Here, this functional group is represented by Cuscuta americana (Convolvulaceae). Contrary to parasitic vines, mistletoes germinate directly upon the branches of their host plants and develop either multiple or solitary haustoria25. The species chosen to illustrate this functional group is Struthanthus martianus (Loranthaceae), which forms various connections with the host branch (Figure 1D). Analysis of solitary mistletoe haustoria using a combination of micro-CT and light microscopy can be found in Teixeira-Costa & Ceccantini17. Finally, obligate root parasites comprise species that germinate on the ground and penetrate the roots of host plants, upon which they are entirely dependent from the earliest growth stages25. These plants are represented here by Scybalium fungiforme (Balanophoraceae), which produce large tuber-like haustoria (Figure 1E).
All plant samples used in this protocol were fixed in a 70% formalin acetic acid alcohol (FAA 70). The fixation upon sampling is crucial for preserving plant tissues, especially if subsequent anatomical analyses are needed. In the case of parasitic plant haustorium, fixation is also essential, as this organ is often primarily composed of non-lignified parenchyma cells20. Detailed protocols for plant tissue fixation, including the preparation of fixative solutions, can be found elsewhere27. On the other hand, to a greater or lesser degree, fixatives can cause alterations of a sample's physical and chemical properties, rendering it unsuitable for specific biomechanical and histochemical analyses. Thus, fresh samples, i.e., non-fixated material collected immediately before preparation, can also be used with this protocol. Details on how to handle fresh samples and troubleshooting suggestions for fixated material are provided in the discussion section.
1. Parasitic plant sample selection
- Collect the entire parasitic plant haustorium, including the attached host stem/root and segments of both proximal and distal ends of the parasitized host organ; the ideal length of each segment is equivalent to double the diameter of the haustorium.
NOTE: For lateral haustoria, include part of the parasite mother stem/root from which the haustorium was formed (Figure 1A,B,D). For endoparasites, collect a segment of the host stem/root in which signs of the parasite are visible (Figure 1B). In the case of terminal connections, the whole plant should be collected (Figure 1E). - Submerge the whole sample in fixative solution (e.g., FAA) in a volumetric proportion of 1:10 (sample: fixative). Leave samples in fixative for at least 1 day, depending on the size of the sample27.
NOTE: Samples can be stored in fixative before scanning or be transferred to a preserving solution (e.g., ethanol 70%). Fresh samples can also be used if the subsequent anatomical analysis is not warranted (see Discussion section). If working with fresh material, set up the apparatus for perfusion of the contrast solution, then collect the sample. The sample should not be allowed to dry out.
2. Application of contrasting solutions
- Chose the application method to be used. Use the vacuum method (step 2.3) for small (Figure 1A) or non-woody (Figure 1B) samples. Use the perfusion method for larger samples, provided it includes a segment of the host stem/root is (Figure 1A,C-E).
- Wear rubber gloves and other appropriate personal protection equipment (e.g., laboratory coat) when manipulating the sample regardless of the approach chosen in the following steps.
CAUTION: All contrasting solutions include heavy metal salts in their composition, and therefore should not be handled without adequate personal protection equipment and under a fume hood. - For the vacuum method, follow the steps mentioned below.
- Choose an appropriate vial and label it. The vial must be large enough to accommodate the sample and the contrasting solution, usually in a proportion of 1:10. Check instructions from the manufacturer to ensure that the vial can withstand low to moderate vacuum.
NOTE: Do not fill the vial to the brim, as the negative pressure (vacuum) can cause the liquid to spill. - Place the sample in the vial with the contrasting solution (1% iodine or 3% phosphotungstate, see Table of Materials). Then place the vial in a vacuum chamber or desiccator connected to a vacuum pump. Remove the lid from the vial, then close the vacuum chamber or desiccator.
- Check that there are no cracks on the vacuum chamber/desiccator and that the vacuum pump has enough oil.
- Close the exhaustion valve of the pump to prevent air from scaping and open the exhaustion valve of the chamber/desiccator to force the air out.
- Turn on the pump and wait until the pressure reaches approximately 20" Hg.
CAUTION: This process is usually fast, so do not leave the vacuum system unattended.
NOTE: While the metric system unit for pressure is Pascal (Pa), pressure gauges in most laboratory vacuum pumps display pressure in inches of mercury ("Hg), pound per square inch (psi), or bars. 20" Hg equals ca. 67.7 Pa, 10 psi, or 0.7 bar. - Close the chamber/desiccator exhaust valve to prevent air from re-entering, then quickly turn off the pump.
- Leave the sample under vacuum for at least 2 h; larger samples require longer for the contrasting solution to penetrate it.
- After the desired period, remove the sample from the contrasting solution to prepare it for scanning.
- Slowly open the chamber/desiccator exhaustion valve to allow air to enter it.
- Wait for the pressure in the chamber/desiccator to be fully exhausted (i.e., pressure gauge reaches near 0), then carefully open it to retrieve the sample.
- Discard the contrasting solution appropriately and keep the sample in preparation for scanning.
- Choose an appropriate vial and label it. The vial must be large enough to accommodate the sample and the contrasting solution, usually in a proportion of 1:10. Check instructions from the manufacturer to ensure that the vial can withstand low to moderate vacuum.
- For the perfusion method, follow the steps mentioned below.
- Select a supply tank for the contrasting solution according to the size of the sample. Use a 50 mL syringe (with no needle or plunger) for small samples (Figure 2A) or a 1 L intravenous medical kit for large samples (Figure 2B).
- Connect one end of a transparent plastic tubing (see Table of Materials) to the supply tank, then connect the other end to a two-way or three-way valve. Connect a second tubing to a different outlet in the valve (Figure 2A,B).
- Secure the supply tank at an elevated position without disassembling the apparatus set up in the previous step.
NOTE: The vertical distance between the tank and the sample will dictate the solution perfusion pressure (Figure 2B). A distance of 20-50 cm is enough for small samples. For large samples, a distance of 1 m is more adequate. - Close the three-way (Figure 2C) or two-way (Figure 2D) valve to prevent liquid from exiting the tubing system, then pour the contrasting solution into the supply tank. If using an intravenous medical kit, fill the bag with the solution and close the valve before securing the apparatus at an elevated position.
- Ensure that no large air bubbles are formed along the tubing system (Figure 2E). If necessary, allow the contrast solution to flow out of the tubing until the bubble is removed. Close the valve again and leave the apparatus in place.
- To prepare the sample for perfusion of the contrast solution, keep it submerged in liquid (water, ethanol, or fixative) and cut off the tip of the proximal end of the host stem/root in the sample (Figure 2F).
- Remove the sample from the liquid in which it was stored and wrap it in a paraffin film to avoid desiccation. Keep the sample nearby and ready to be connected to the apparatus.
- Carefully open the valve to allow the contrasting solution to flow slowly and fill the plastic tubing connected to the tank while holding the open end of the system at a slightly elevated position to prevent the contrasting solution from spilling. Again, ensure that no large air bubbles form along the tube.
- Connect the proximal end of the host stem/root in the sample to the open end of the tubing system (Figure 2B, magnified region). Avoid introducing air bubbles into the system during this step. If necessary, disconnect the sample from the apparatus and remove air bubbles from the system by allowing the solution to flow.
- While keeping the sample connected to the apparatus, place it inside a container to avoid leakage of the contrasting solution into the area where the experiment was set up. Use tubes of different diameters, plastic zip-ties, and valve adaptors (Figure 2G) to ensure all connections in the apparatus are well-fit, accommodate host branches of various sizes, and that the solution is not leaking out of the tubing system (Figure 2B, magnified region).
- Let the solution perfuse the sample for at least 2 h, or until the solution accumulates inside the container.
- Close the valve and carefully disconnect the sample from the apparatus. Drain the remainder of the solution into the container and dispose of it appropriately.
- Remove the paraffin film from the sample in preparation for scanning.
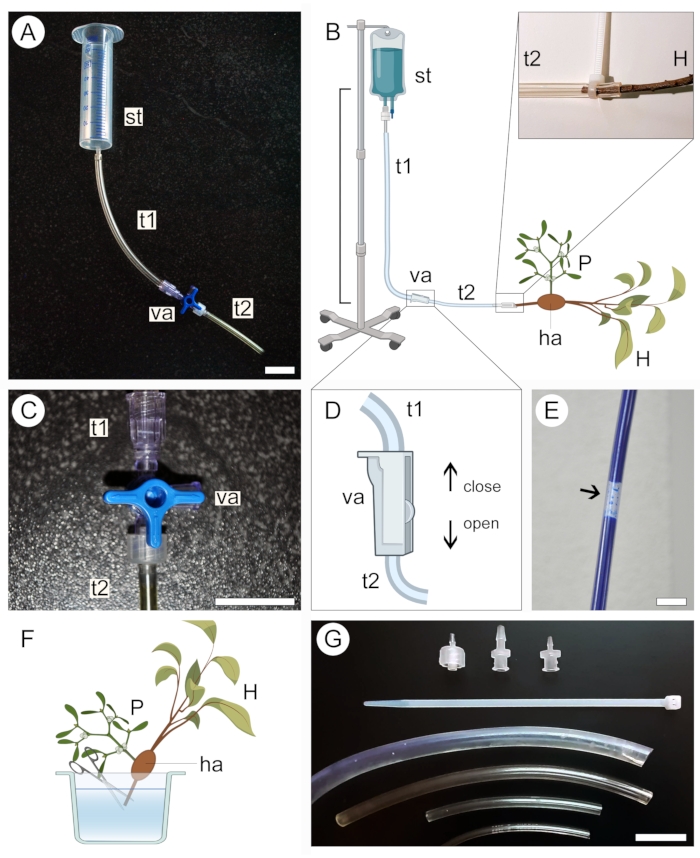
Figure 2: Perfusion approach for contrast application. Small (A) and large (B) versions of the perfusion apparatus include a supply tank (st) and two plastic tubes (t1 and t2) connected by a valve (va). The proximal end of the host stem (H) bearing a parasite (P) attached to it via the haustorium (ha) is connected to the open end of the system (B, expanded). A three-way (C) or a two-way (D) valve is used to help prevent the formation of air bubbles inside the tubing system, which block the passage of contrasting solution (E). The tip of the proximal end of the host stem (H) is cut underwater to allow passage of the contrast solution (F). Zip-ties, valve adaptors, and tubing of different diameters help secure tighter connections and avoid leakage in the system (G). Figures 2B, D and F were created with BioRender. Scale bars = 2 cm. Please click here to view a larger version of this figure.
3. Sample preparation and mounting
- Wash the sample by submerging it in water for 2 min.
CAUTION: Do not wash samples in the sink, as all contrasting solutions include heavy metal salts in their composition. Consider the water used for washing as a diluted contrasting solution and dispose of it appropriately. - Place the sample in a paper towel at room temperature to allow excess water to evaporate for 2-5 min depending on the size of the sample. Alternatively, dry the sample slightly with the aid of a paper towel. Do not allow the sample to dry out completely.
- Wrap the sample in a paraffin film by stretching it to a thin layer. Avoid folding the paraffin film on top of the sample.
- Mount the wrapped sample onto a sample holder, keeping it stable and in position while it rotates during the scanning. Use adhesive tape, low-density foam, pipette tips, and/or clear plastic containers to secure the sample in place.
- Scan the sample and analyze the images following specific protocols and guidelines established for the micro-CT system available.
The haustorium of parasitic plants is a complex organ comprising different tissues and cell types that intertwine and connect with the tissues of another plant, used as a host20. Micro-CT scanning can be leveraged to better understand this complex structure in a non-destructive and three-dimensional way when analyzing both small (Figure 1A-C) and large (Figure 1D,E) haustoria. To do this, contrasting solutions can be applied into the parasite-host interface, thus making it possible to analyze samples that would otherwise have low and homogenous X-rays absorption. The two simple approaches described in this protocol rely on pressurizing the contrasting solution to accelerate penetration through the sample. As expected, the protocols described here work for different micro-CT systems. However, scanning settings and parameters differ depending on the system and the analyzed sample (Table 1).
| Functional group | Euphytoid parasite | Endoparasite | Parasitic vine | Mistletoe (before/after contrast) | Obligate root parasite |
| Species (Family) | Pyrularia pubera | Viscum minimum | Cuscuta americana | Struthanthus martianus | Scybalium fungiforme |
| Family | Santalaceae | Viscaceae | Convolvulaceae | Loranthaceae | Balanophoraceae |
| Contrast solution | 3% phosphotungstate | 1% iodine | 1% iodine | 1% iodine | 0.2% lead nitrate |
| Application method | vacuum | vacuum | perfusion | perfusion | perfusion |
| Micro-CT system | Zeiss Versa 620 | Nikon X-Tek HMXST225 | Bruker Skyscan 1176 | Bruker Skyscan 1176 | Bruker Skyscan 1176 |
| Projections | 4500 | 3140 | 610 | 1200 / 2400 | 5300 |
| Frames | 1 | 1 | 3 (averaged) | 3 / 3 (averaged) | 3 (averaged) |
| Exposure (ms) | 5000 | 1000 | 680 | 1600 / 830 | 750 |
| Filter (mm) | none | Al 0.25 | Al 1.0 | Al 1.0 / Al 1.0 | none |
| Voltage (kV) | 60 | 85 | 35 | 90 / 45 | 40 |
| Current (μA) | 108 | 80 | 375 | 180 / 390 | 600 |
Table 1: Scanning settings and parameters for the analyzed samples.
Following the first approach for contrast application (step 2.3), vacuum was used to perfuse the haustorium of the euphytoid parasite P. pubera with a 3% phosphotungstate solution for two hours. The sample was then scanned in a 3D X-ray microscope (XRM) system (see Table of Materials), achieving high image resolution via secondary optical magnification4. XRM images were observed to be as effective as anatomical sections analyzed under a light microscope to analyze tissue organization and topology at the parasite-host interface (Figure 3). The use of phosphotungstate highlights the abundance of parenchyma tissue, which appears in a bright white tone due to the high absorption of the contrasting agent. Vessels appear in a dark gray tone due to low absorption of contrast. Based on this color difference, it is possible to observe the abundance of parenchyma surrounding the vascular core of the haustorium (Figure 3). The intricate vessels and thin parenchyma strands of the vascular core itself are also observed, especially in longitudinal sections (Figures 3A-C). In cross-sections, the vascular core is easily observed as two vascular strands separated by parenchyma (Figures 3D,E).
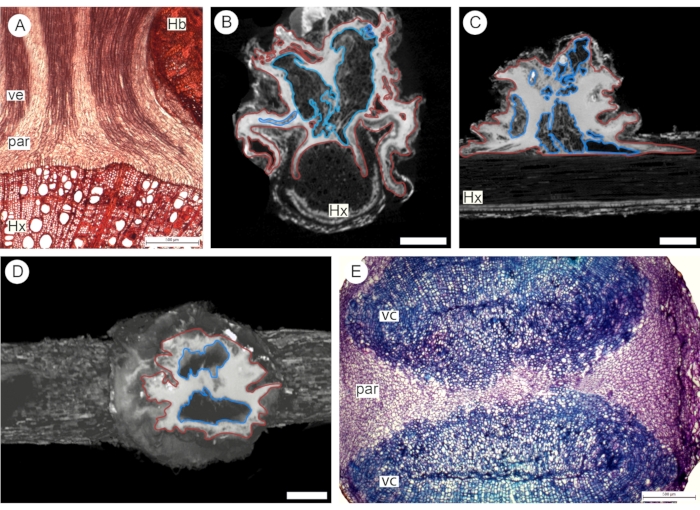
Figure 3: Euphytoid parasite haustorium. Longitudinal (A-C) and transversal (D-E) sections through the haustorium were observed under a light microscope (A,E) and via 3D X-ray microscopy following application of 3% phosphotungstate solution using vacuum (B-D). Red outlines indicate parenchyma tissue (par), and blue outlines indicate the vascular core (vc). Hx: host xylem. Hb: host bark. Scale bars = 500 µm (A, E) and 2.5 cm (B-D; voxel size = 2.8382 µm). Please click here to view a larger version of this figure.
The same approach was used to introduce contrast into the stem of a succulent host plant (Euphorbia polygona, Cactaceae) parasitized by V. minimum. Here, 1% iodine solution was chosen as a contrasting agent based on preliminary histochemical analysis, which showed that parenchyma cells of the endoparasite store carbohydrates in the form of starch (Figure 4A). At the same time, cells in the host cortex do not store a detectable amount of starch (Figure 4B-D). After contrast application, the sample was then scanned using a Nikon X-Tek micro-CT system (see Table of Materials). The difference in iodine absorption allowed the detection of the intricate web of cortical strands formed by the endoparasite within the host body (Figure 4E). Following this methodology, cortical strands were observed to concentrate around the host vascular center and eventually branch towards the periphery of the host stem when associated with the emersion of a parasite flower (Figures 4F,G).
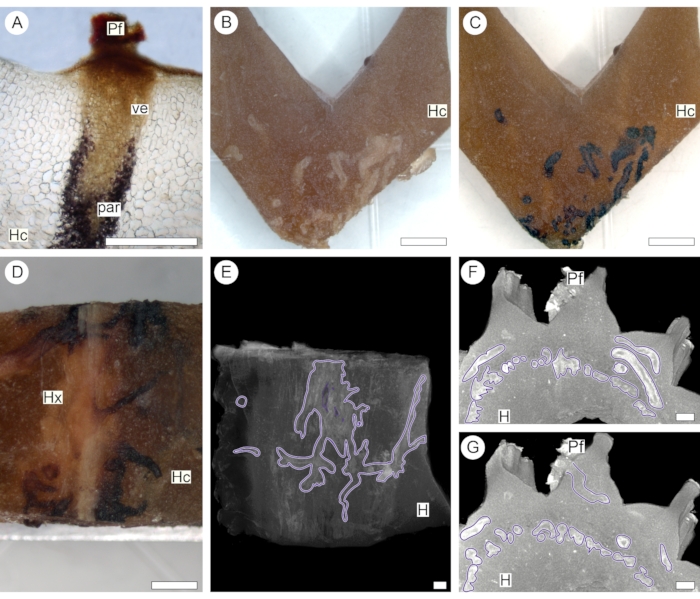
Figure 4: Endoparasite tissue network. Anatomical (A) and macro (B-D) sections show reaction with 1% iodine solution indicating that starch (stained in black) is present in the parenchyma (par) associated with the vessels (ve) of the parasite. Because starch is absent in the host cortex (Hc) and xylem (Hx), 1% iodine solutions were selectively used to enhance the contrast of parasite cortical strand tissues (purple outlines, E-G), seen against the host background (H). The presence of a flower (Pf) confirms that the stained tissue is part of the endoparasite structure (A,F,G). Scale bars = 0.25 cm. Please click here to view a larger version of this figure.
In the second approach described here (step 2.4), a supply tank containing the contrasting solution is elevated from the level in which the sample is placed, thus using gravity to drive the passage of the solution through the sample. After contrast perfusion, scanning was carried out using a Bruker Skycan micro-CT system (see Table of Materials). Results obtained for C. americana (Figures 5A,B) and S. martianus (Figures 5C-G) help illustrate the convenience of this approach for both small and large samples, respectively. Comparing samples scanned before (Figure 5C,E) and after (Figure 5D,F) perfusion of 1% iodine solution stresses the importance of contrast application even in woody haustorium samples. It is noteworthy that in the associations between both C. americana and S. martianus (Figure 5) and their respective hosts, parasites have little to no starch content in their tissues. This explains the different results described for the endoparasite V. minimum (Figure 4), in which the same method and type of contrast solution were used.
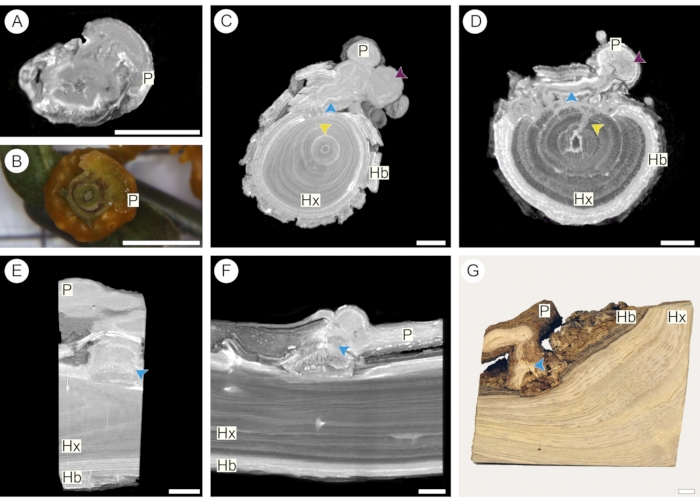
Figure 5: Parasitic vine and mistletoe haustorium. Longitudinal sections through the haustorium of a parasitic vine (A,B) and a mistletoe (C-G). Comparison between scan and macro section of fresh material shows that the intricate haustorium structure is well captured in micro-CT scans (A,B). Comparison between fixated samples before (C,E) and after (D,F) the perfusion of 1% iodine solution show that contrast is enhanced even in lignified samples, facilitating the observation of parasites (P) structures such as vessels in the epicortical root (pink arrowheads), vascular strands (blue arrowheads), and sinker (yellow arrowhead). Vessels and annual rings of the host xylem (Hx) and starch in the host bark (Hb) are also more easily observed. Scale bars = 1 cm. Please click here to view a larger version of this figure.
A final result obtained with the perfusion protocol described here is the possibility to detect vascular connections between parasites and host plants. This was achieved by perfusing a 0.2% lead nitrate solution through a segment of the host root around which the tuber of the obligate root parasite S. fungiforme had developed. After contrast application, scanning was also carried out using a Bruker Skyscan micro-CT system (see Table of Materials). Sequential projections show that vessels in the host root bifurcate into the parasite tuber (Figure 6A-C). Following the analysis of these results, the same sample was cut into a smaller sub-sample, prepared for anatomical study, and analyzed using light microscopy. Serial sectioning of this sub-sample confirms that the xylem continuity between the two plants is formed by vessel-to-vessel connection via perforation plates (Figure 6D-F).
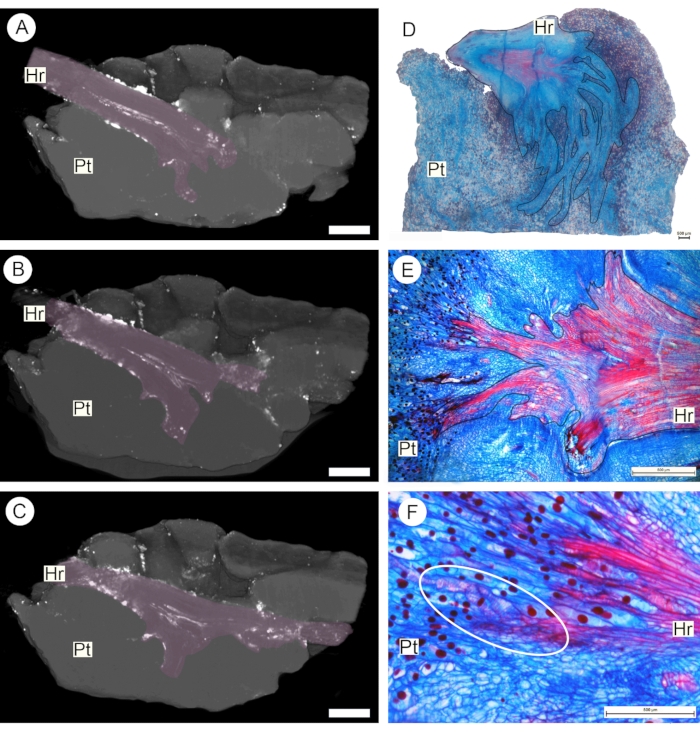
Figure 6: Obligate root parasite haustorium. Longitudinal sections through the haustorium as observed via micro-CT scanning (A-C) and anatomical sections observed under the light microscope (D-F). The accumulation of 0.2% lead nitrate solution within the vascular system of the host root (Hr, pink outline) allows observation of host vessels branching within the parasite tube (Pt) and the detection of direct vascular connection between the two plants (dashed white outline). Scale bars = 1 cm (A-C) and 500 µm (D-F). Please click here to view a larger version of this figure.
