When using density gradient to purify neutrophils, it is critical for the interface between the blood and the density gradient medium to be as sharp as possible, and that a distinct layer separation remains after centrifugation (step 1.4). Following RBC lysis, the buffer should be a clear red and not turbid (step 3.3). If the preparation is cloudy, a second round of lysis (step 3) may be required, although this may affect neutrophil survival (Figure 1). Following lysis, low-speed centrifugation (200 × g) is recommended when purity is prioritized, as it reduces platelet contamination considerably. However, high-speed centrifugation (400 × g) increases the yield at the expense of purity (step 3.4, Figure 2). After neutrophil isolation, fluorescence-activated cell sorting can be used to assess isolation purity (step 4.1) and should be chosen over microscopy. Although the FSC/SSC distribution of cells alone provides an estimate of the cell isolation quality (Figure 3A), the use of specific cell markers should be preferred. In this case, the most common contaminating cell populations are stained with specific antibodies together with CD66b, specifically expressed on granulocytes. CD45 staining is used to distinguish leukocytes (CD45+) and red blood cells and platelets (CD45-).
Other contaminants include lymphocytes (CD3+ or CD19+, Figure 3F), monocytes (CD14+, Figure 3D), and eosinophils (CD193+, Figure 3G). CD11b is an integrin expressed on the myeloid lineage; neutrophils and monocytes are CD11b+, whereas lymphocytes are CD11b- (Figure 3C). As neutrophil activation can affect downstream experiments, the expression of CD62L should be assessed; neutrophils become CD62L- once activated (step 4.1). The peptide fMLP can be used as a positive control for CD62L shedding (Figure 4). It is also important to evaluate the health of neutrophils before performing assays; neutrophils have a relatively short half-life, and activation can further shorten it (step 4.2). A standard Annexin V and PI staining can give information on the live/dead status of the neutrophil culture at designated times (Figure 5).

Figure 1: Density gradient medium-based separation of granulocytes. (A) Before and (B) after centrifugation. Note the sharp interface between the blood and the density gradient medium layers. (C) Sedimentation of the pellet in B resuspended (1:1) in 5% FBS/HBSS and 3% Dextran-0.9% NaCl. (D) PMN pellet after lysis of the residual RBCs in the supernatant of (C) with H2O. Abbreviations: PBMC = peripheral blood mononuclear cell; PMN = neutrophil; RBC = red blood cell; FBS = fetal bovine serum; HBSS = Hank's Balanced Salt Solution. Please click here to view a larger version of this figure.
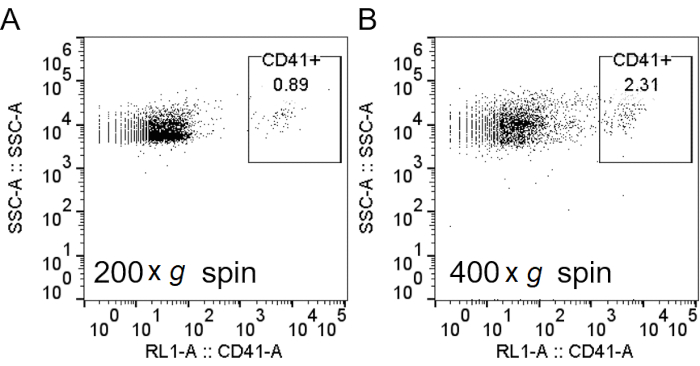
Figure 2: Spinning at lower speeds after RBC lysis decreased platelet contamination. After lysis of RBCs with H2O, cells were spun down at 200 × g (A) or 400 × g (B). Neutrophils were stained, and flow cytometry analysis was performed, as described in Figure 3, with the addition of anti-CD41 to label the platelets. Abbreviations: RBC = red blood cell; CD41 = cluster of differentiation 41; SSC-A = side scatter-Area. Please click here to view a larger version of this figure.
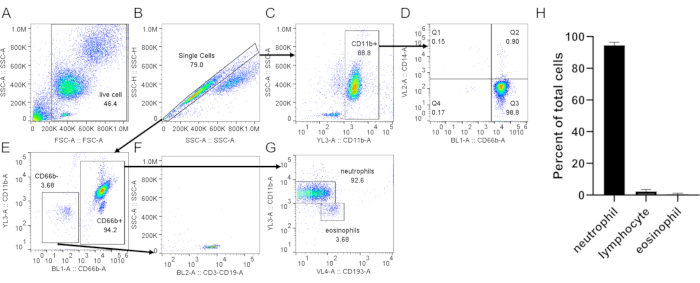
Figure 3: Assessment of neutrophils isolated from buffy coat. Isolated neutrophils were stained with anti-CD66b, anti-CD11b, anti-CD14, and anti-CD193 following a standard protocol. Single cells (B) were gated from total cells (A). (C) CD11b+ cells were gated from single cells. (D) Dot plot showing neutrophils (CD66b+, CD14 low/-) and a very low monocyte contamination (CD66b–, CD14+, Q1). (E) CD66b– and CD66b+ cells were gated. (F) CD66b– cells were positive for lymphocyte markers (CD3, CD19). (G) Expression of CD11b and CD193 in CD66b+ cells, showing distinct neutrophil (CD66b+, CD11b+, CD193–) and eosinophil (CD66b+, CD11b–, CD193+) populations. In the representative result shown here, neutrophil purity is ~93% with ~3.7% lymphocyte and ~3.7% eosinophil contamination. (H) Quantification of neutrophil purity after purification. Data are compiled from 5 individual trials and presented as mean ± SD. Abbreviations: CD = cluster of differentiation; SSC-A = SSC-A = side scatter-Area; FSC-A = forward scatter-Area. Please click here to view a larger version of this figure.
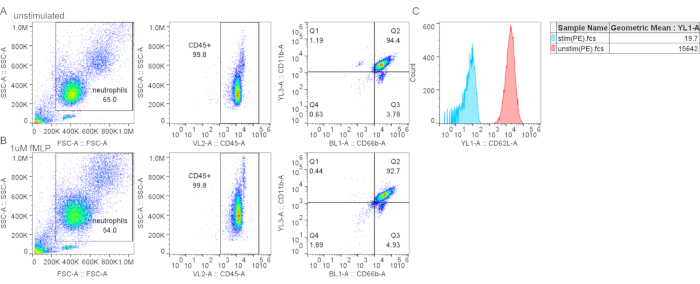
Figure 4: Gradient purification did not cause CD62L shedding. (A–B) Control (A) or fMLP-stimulated neutrophils (B) (1 mM fMLP for 15 min at 37 °C) were stained with neutrophil markers and CD62L. (C) Fluorescent mean intensity for CD62L is decreased in fMLP-treated cells, indicating CD62L shedding and neutrophil activation. Abbreviations: CD26L = L-selectin; fMLP = N-Formylmethionyl-leucyl-phenylalanine; SSC-A = SSC-A = side scatter-Area; FSC-A = forward scatter-Area; PE = phycoerythrin. Please click here to view a larger version of this figure.
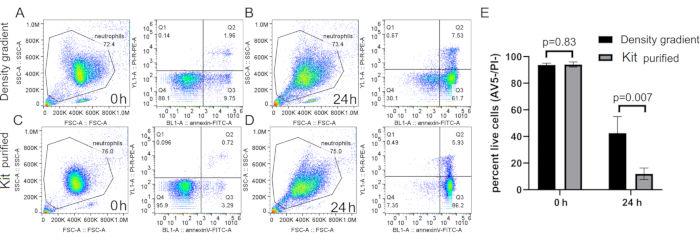
Figure 5: Spontaneous death of neutrophils purified by density gradient or neutrophil isolation kit. Neutrophils were purified with density gradient (A and B) or commercial microbeads (C and D) and cultured for 0 h (A and C) or 24 h (B and D) in RPMI-10% FCS. Cells were stained using Annexin V and PI at respective time points following a standard protocol. (E) Quantification of purified neutrophil spontaneous death. n=5, mean ± SD. Abbreviations: SSC-A = SSC-A = side scatter-Area; FSC-A = forward scatter-Area; PI = propidium iodide; FCS = fetal calf serum; FITC = fluorescein isothiocyanate; AV5 = Annexin V. Please click here to view a larger version of this figure.
