Each Japanese plum flower bud contains an inflorescence with 1–3 flowers. As in other stone fruit species, each flower is made up of four whorls: carpel, stamens, petals, and sepals, which are fused forming a cup at the base of the flower. Flower structures are smaller than other stone fruits, with a short and fragile pistil surrounded by the stamens that contain a small amount of pollen grains. At full bloom, the flowers of each inflorescence appear separated on short stalks, showing the white petals forming a balloon surrounded by the green sepals (stage D, 57 BBCH) (Figure 1A) in the days before anthesis. The flower is fully open at anthesis, showing the anthers and the pistil (stage F, 65 BBCH) (Figure 2A). Like other temperate Prunus spp., the flower buds open first, and the leaf buds sprout several days later. This makes blooming trees look spectacular showing a great number of flowers but no leaves.
Hand-pollinations in Japanese plum required the collection of flowers at the balloon stage previous to anthesis (Figure 1A), in which the pistil and stamens are nearly mature, but the anthers are still undehisced. This stage prevents the arrival of insects carrying external pollen because the petals are still closed. Although the number of flowers is much higher than in other Prunus species, most of them (85%–95%) are not able to set fruit. As a result, using a high number of flowers for pollination experiments is mandatory. The undehisced anthers could be easily separated from the flower in the laboratory and extended on a piece of paper (Figure 1B), where the anthers are dehisced after 24 h at room temperature, showing the pollen grains (Figure 1C). Then, the pollen grains were easily sieved through a fine mesh (Figure 1D), and could be used immediately or stored until use, both for field (Figure 2A–D) and/or laboratory pollinations (Figure 2E,F).
The monitoring of fruit drop of hand-pollinated flowers after emasculation (Figure 2A,B) or supplementary pollination of non-emasculated flowers (Figure 2C,D) showed clear differences between treatments. Most flowers dropped 2–3 weeks after pollination in both treatments. All self-pollinated flowers dropped. However, 4% of self-pollinated flowers from the supplementary pollination treatment remained in the tree until harvest, indicating that this cultivar (Rubirosa) behaves as self-compatible (Figure 3). The behavior of cross-pollinated flowers used as control also varied between treatments. Cross-pollinated flowers with supplementary pollination dropped until 3 weeks after pollination, resulting in 7% of fruit set. However, all emasculated cross-pollinated flowers dropped coinciding with the drop of all self-pollinated flowers.
For hand pollinations in the laboratory, the pistils were placed in wet florist foam after a fresh cut on the base of each flower pedicel underwater (Figure 2E) and pollinated 24 h later with the help of a fine brush (Figure 2F). In self-pollinated flowers, self-compatible cultivars showed at least one pollen tube reaching the base of the style in most of the pistils examined, while in self-incompatible cultivars, pollen tube growth was arrested in the upper style (Table 2).
In vitro pollen germination differed significantly between cultivars (Table 3). Good pollen viability was considered when more than 20% of pollen grains showed a pollen tube longer than its length after 24 h in the culture medium (Figure 4A). However, when most of pollen grains did not germinate (Figure 4B), the cultivar was considered male sterile and not suitable as a pollinator.
Like in other Prunus, the pistil is made up of three structures: stigma, style, and ovary. The ovary has two ovules, and at least one of them should be fertilized for fruiting. During pollination, pollen grains are transferred to the stigma, where germination occurred within 24 h (Figure 4C). Each germinating pollen grain produced a pollen tube, which grew through the pistil structures. In self-incompatible cultivars in which the pollen grains have an S allele that coincides with one of the two S alleles of the pistil, the pollen tube stopped growing in the upper third of the style (Figure 4D), preventing the arrival to the ovary and the subsequent fertilization. However, when S allele of the pollen grain is different with that of the pistil, the pollen tube could grow through the style (Figure 4E), reach the ovary (Figure 4F) and fertilize an ovule.
PCR analysis (Table 1) was carried out using primers from conserved regions of S-RNase of sweet cherry and Japanese plum (Figure 5). The primer set used, PruC2-PCER and PruT2-PCER (See Table of Materials), allowed to determine the size of both S-alleles in each cultivar. The amplified fragments were run in agarose gel by electrophoresis, and seven different S-alleles (Sa, Sb, Sc, Se, Sf, Sh, Sk) were identified in the cultivars analyzed. The fragment sizes ranged between 393 and 1,580 bp using PruC2-PCER (Figure 5) and between 820 and 1,993 bp using PruT2-PCER. Six S-genotypes were identified (SaSb, SbSc, SbSf, ScSh, SeSh, SfSk).
The phenology of each cultivar allowed to calculate the length of the flowering period for a total period of four years, considering full bloom when most flowers were at stage F, stage 65 BBCH. Flowering in orchard conditions (Figure 6) allowed the comparison of flowering times between cultivars and years, and the determination of which cultivars are coincident at flowering time in each year (Figure 7).

Figure 1: Pollen extraction in Japanese plum. (A) Flower buds at balloon stage D, according to Baggiolini24, stage 57 of the BBCH scale. (B) Removal of undehisced anthers from the flower. (C) Dehisced anthers showing the pollen grains. (D) Sieve of pollen grains from anthers using a fine mesh. Please click here to view a larger version of this figure.

Figure 2: Hand-pollination experiments to determine self-(in)compatibility in Japanese plum. Field pollinations: (A) Flowering in Japanese plum, with flowers at anthesis and at balloon stage. (B) Flowers at balloon stage D, according to Baggiolini24, stage 57 of the BBCH scale after emasculation. (C) Caged tree to avoid the arrival of insects. (D) Supplementary hand-pollination of non-emasculated flowers. Laboratory pollinations: (E) Cut on the base of pedicel underwater and emasculated flowers placed on soaked foam. (F) Hand-pollination of the pistils with a fine paintbrush. Please click here to view a larger version of this figure.
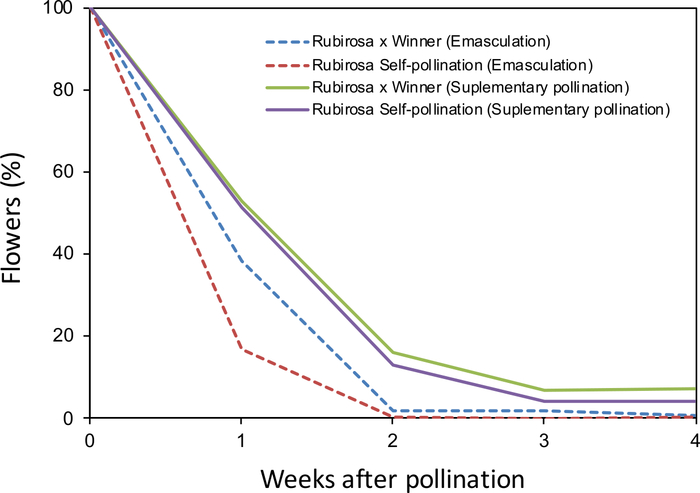
Figure 3: Fruit drop in Japanese plum as affected by different pollination treatments. Self- and cross-pollination in emasculated and non-emasculated flowers. Percentage of flowers and developing fruits from the original number of remaining flowers in the tree during the 4 weeks after pollination in 'Rubirosa'. Please click here to view a larger version of this figure.
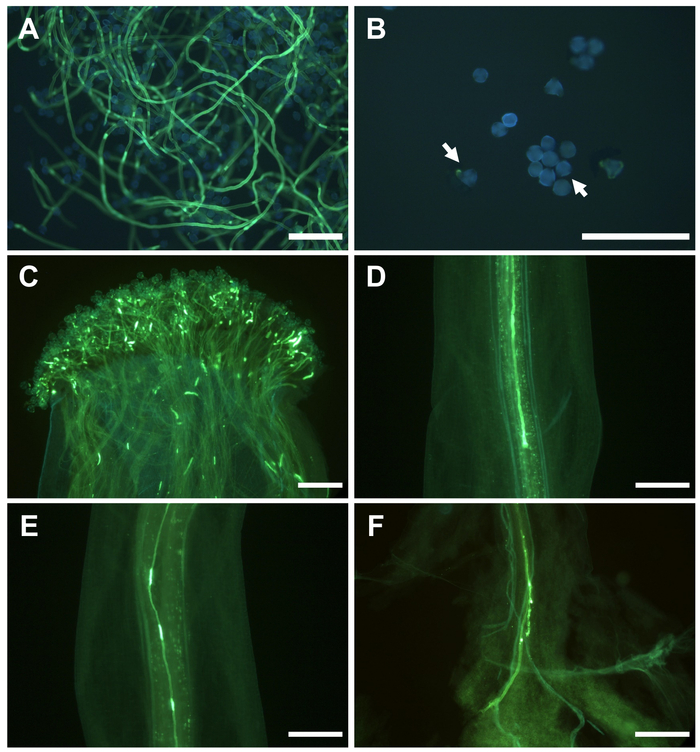
Figure 4: Pollen germination and pollen tube growth in self-pollinated flowers in Japanese plum. (A) In vitro pollen germination. (B) Non-germinated pollen grains (arrows) in vitro. (C) Pollen grain germination on the stigma surface. (D) Pollen tube arrested in the upper third of the style. (E) Pollen tube growing along the style. (F) Pollen tubes at the base of the style. Scale bars, 200 μm. Please click here to view a larger version of this figure.
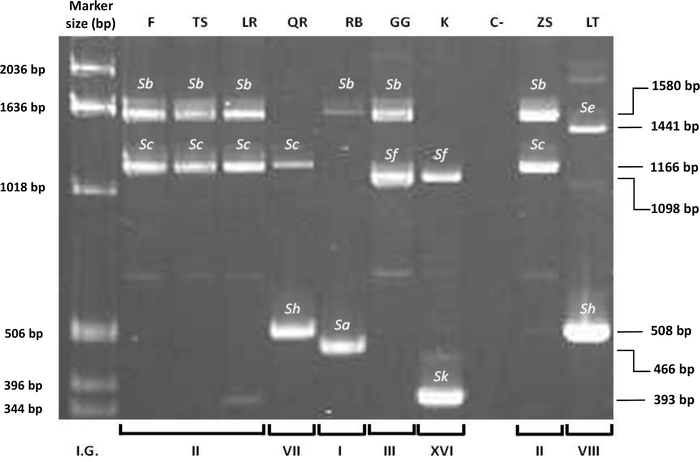
Figure 5: PCR amplification using primer set PruC2-PCER of nine Japanese plum cultivars. Identification of seven S-alleles (Sa, Sb, Sc, Se, Sf, Sh, Sk) and six S-genotypes (SaSb, SbSc, SbSf, ScSh, SeSh, SfSk). Incompatibility Group (I.G.3), ‘Fortune’ (F), ‘TC Sun’ (TS), ‘Laroda’ (LR), ‘Queen Rosa (QR), ‘Red Beaut’ (RB), ‘Golden Globe’ (GG), ‘Kelsey’ (K), Negative control (distilled water) (C-), ‘Zanzi Sun’ (ZS), ‘Laetitia’ (LT). 1 kb: Size standard. Please click here to view a larger version of this figure.

Figure 6: Monitoring of phenology of Japanese plum. Two Japanese plum cultivars with flowers in different phenological stage. Stage C24, stage 55 of BBCH scale (left) and Stage F24, stage 65 of BBCH scale (right). Please click here to view a larger version of this figure.
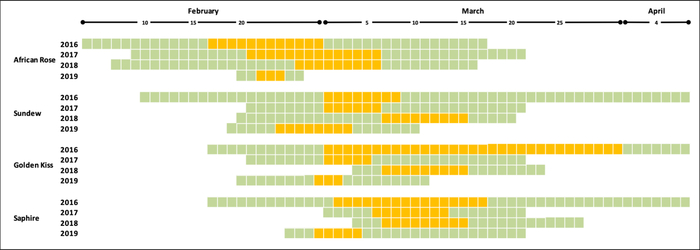
Figure 7: Flowering time in four Japanese plum cultivars over 4 years. Period from the first to the last open flowers. Yellow cells indicate the days of full bloom in which most flowers were open. Please click here to view a larger version of this figure.
| Reagents | Volume per one reaction (μL) | |
| H2O | 11.50 | |
| 10X Buffer with 20 mM MgCl2 | 2.80 | |
| dNTP mix , 10 mM each | 0.80 | |
| Primer forward | 0.40 | |
| PruC2 (5'-CTATGGCCAAGTAATTATTCAAACC-3')40 or | ||
| PruT2 (5'- TSTTSTTGSTTTTGCTTTCTT-3')40 | ||
| Primer reverse | 0.40 | |
| PCER (5'-TGTTTGTTCCATTCGCCTTCCC-3')41 | ||
| DNA template | 4.00 | |
| Taq DNA polymerase, 500 U | 0.09 | |
| Final volume | 20.0 | |
Table 1: Reaction conditions used in this protocol.
| Cultivars (S– genotype) | Number of pistils examined | Germinated pollen grains on stigma (%) | Pistils with pollen tubes at the base of style (%) | Pollen tubes at the base of style (mean number) | Self/cross -compatibility reaction |
| TC Sun (SbSc) × Larry Ann (SbSh) | 22 | 92.3 | 27 | 1.2 | + |
| TC Sun (SbSc) × Blackamber (SbSc) | 10 | 74.8 | 0 | 0.0 | – |
| TC Sun (SbSc) self-pollination | 44 | 78.3 | 7 | 1.0 | + |
| Golden Plum (SbSc) × Black Star (SeSf) | 11 | 64.7 | 36 | 1.5 | + |
| Golden Plum (SbSc) × TC sun (SbSc) | 11 | 98.4 | 0 | 0.0 | – |
| Golden Plum (ShSk) self-pollination | 38 | 85.2 | 0 | 0.0 | – |
Table 2: Pollen germination and pollen tube growth through the style for two Japanese plum cultivars after self- and cross-pollinations. Number of hand-pollinated pistils examined, percentage of germinated pollen grains on stigma, percentage of pistils with pollen tubes at the base of the style, mean number of pollen tubes at the base of the style, Self- or cross-incompatible (-) and self- or cross-compatible (+).
| Cultivar | Germination (%) | SD* |
| Earlemoon | 50 | 3.3 |
| Earliqueen | 30 | 4.2 |
| Eldorado | 10 | 1.3 |
| Friar | 52 | 2.0 |
| Golden Japan | 17 | 3.8 |
| Golden Plumza | 18 | 3.3 |
| Laroda | 46 | 3.6 |
| Larry Ann | 20 | 5.8 |
| Methley | 2 | 0.8 |
| Owen T | 16 | 0.3 |
| Primetime | 20 | 3.3 |
| Queen Rosa | 42 | 3.9 |
| Royal Diamond | 19 | 2.2 |
| Santa Rosa | 36 | 2.0 |
| TC Sun | 49 | 3.1 |
| *SD = Standard deviation | ||
Table 3: Percentage of in vitro pollen germination of 15 Japanese plum cultivars. Mean ± SD of six replicates.
