A typical response to the positive control (10-3 M L-serine; Figure 4A) and negative control (blank; Figure 4B) recorded from the olfactory nerve of a seabream is shown in Figure 4. In the presence of the stimulus (black horizontal bar; in the olfactory cavity, in contact with the olfactory epithelium), note the rapid increase in activity (reflected in the upward deflection of the integrated signal) to a peak within about one second of stimulus onset, followed by a period of accommodation (while the stimulus is still present), and a return to baseline activity once the stimulus has ended. The absolute amplitude of the response is highly dependent on electrode position; if a low amplitude response is recorded, try changing the electrode positions. A slower rise to peak activity may be due to the tube carrying the stimulus-containing water to the olfactory epithelium being placed too far away from the epithelium; try moving the nose-tube closer to (but not touching) the epithelium. Note that, in contrast, the blank evokes little or no response. A significant positive response (i.e., increase in activity) to the blank may indicate contamination of the water used to make the dilutions of the stimuli; making fresh dilutions with clean water (and glassware) should resolve this. If not, a more thorough cleaning of the water system (including activated charcoal filters) may be necessary. A negative response (i.e., decrease in activity) may indicate a slight change in flow rate when the valve is switched due to, for example, a blockage in the valve.
A typical concentration-response curve (plotted semi-logarithmically), in this case to L-leucine (10-7 M to 10-3 M), is shown in Figure 5A. Note that increasing concentrations of the odorant evoke increasingly large increases in activity, and therefore, in the amplitude of the integrated responses. A plot of the normalized data, and their corresponding linear regression, is shown in Figure 5B. The estimated threshold of detection can be calculated from the value of x when y = 0.1761 (i.e., log1.5; where N = 0). In this case, this value is -7.48; that is, the calculated threshold for L-leucine in this fish is 10-7.48 M. The exponent α can similarly be estimated from linear regression of the normalized data on a log-log plot; logN = αlog[odorant] + constant. The factor γ then gives the increase in odorant concentration required to increase the response amplitude by one log unit; that is, it is an estimate of the steepness of the concentration-response curve17. In this example, α = 0.277 and γ = 3.61; therefore, to increase the response amplitude ten-fold (i.e., one log unit; log10 = 1), the stimulus concentration needs to be increased by 103.61-fold (4,074-fold).
A typical sigmoidal concentration-response curve (Figure 6A) when plotted semi-logarithmically, in this case to L-glutamine, is shown in Figure 6B. A similar concentration-dependent increase in response amplitude is seen; however, at the higher concentrations, this increase becomes less so that the response amplitude reaches a maximum (Nmax). This allows the data to be fitted to a three parameter Hill equation:
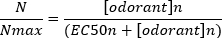
This way, the EC50 (the odorant concentration at which a 50% maximal response is evoked) and the Hill co-efficient (a measure of the steepness of the slope of the linear part of the sigmoidal curve) can be calculated.
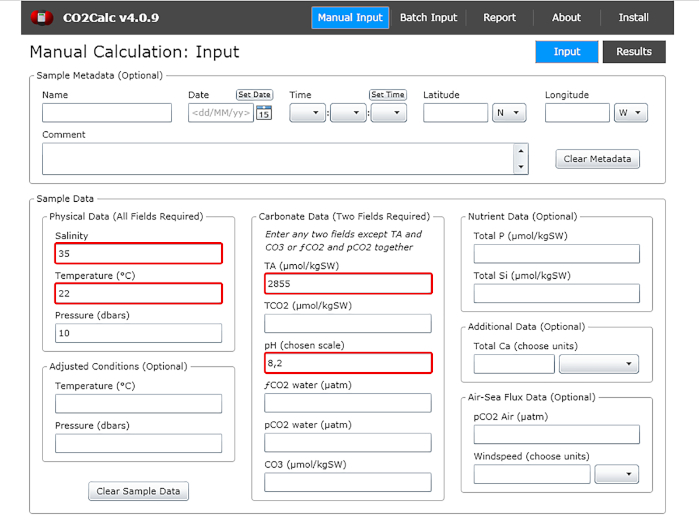
Figure 1: Software screenshot showing the input window from the program CO2Calc. Highlighted (red boxes) are the fields required for carbonate parameter calculation. Please click here to view a larger version of this figure.
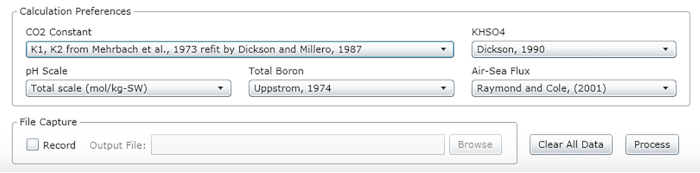
Figure 2: Software screenshot showing the input window for the appropriate constants, units and scales. Values shown are recommended for conditions under which the described experiments were carried out; they may change. Please click here to view a larger version of this figure.
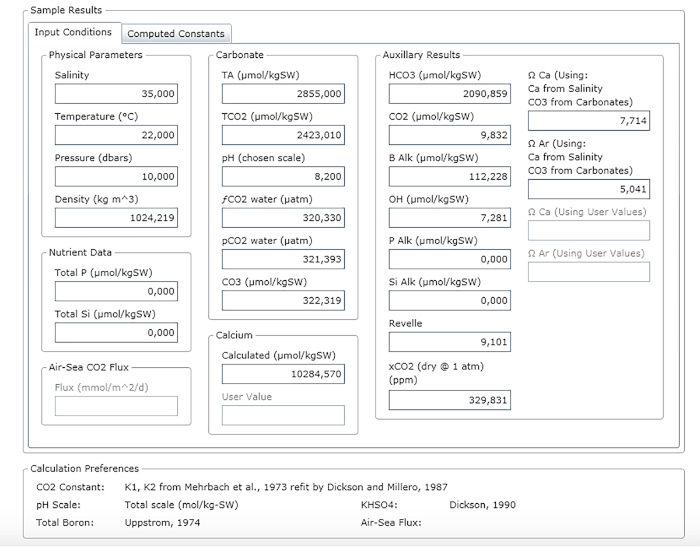
Figure 3: Software screenshot showing the results window. Please click here to view a larger version of this figure.
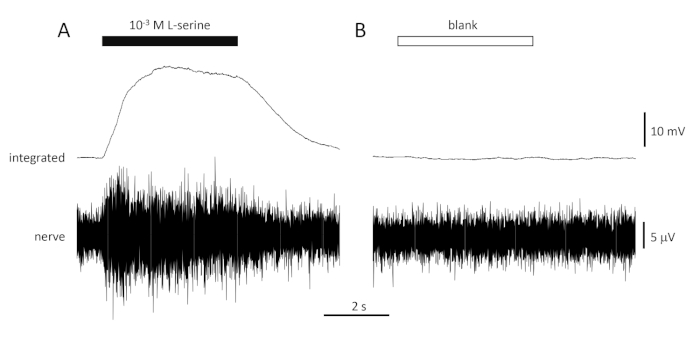
Figure 4: Typical multi-unit responses recorded extracellularly from the olfactory nerve of seabream in vivo in response to 10-3 M L-serine (A) and blank (B). Upper traces show the integrated responses and lower traces show the raw (nerve) signal. Stimuli were applied to the olfactory epithelium (horizontal bars). Note the rapid increase in activity during the 1 s of exposure, a peak in activity, followed by a period of accommodation (while the odorant was still delivered to the epithelium) and a return to baseline levels once the odorant delivery has ceased. Little or no increase in activity is seen following stimulation with water treated the same way as odorant dilutions, with the exception of adding any odorant (blank). Please click here to view a larger version of this figure.
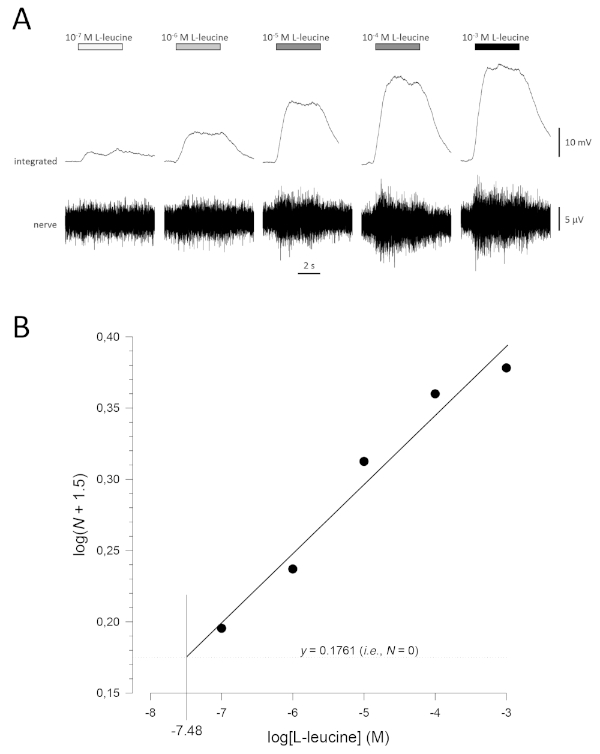
Figure 5: Typical concentration-response curve for L-leucine recorded extracellularly from the olfactory nerve in vivo. (A) As the concentration of L-leucine applied to the olfactory epithelium (horizontal bars) increases from 10-7 M to 10-3 M, a concomitant increase in activity is seen in the nerve. Upper traces show the integrated responses and lower traces show the raw (nerve) signal. (B) Linear regression (R2 = 0.97) of normalized data plotted semi-logarithmically to calculate threshold of detection as the value for log[L-leucine] when log(N + 1.5) = 0.1761 (i.e., where N = 0). In this example, this value is -7.48; the estimated threshold of detection for L-leucine in this fish is therefore 10-7.48 M. Please click here to view a larger version of this figure.
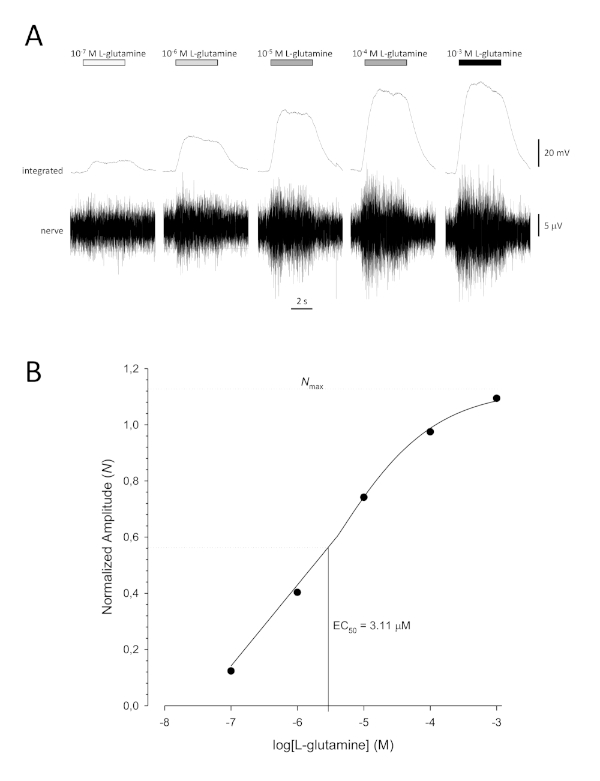
Figure 6: Typical concentration-response curve for L-glutamine recorded extracellularly from the olfactory nerve in vivo. (A) As the concentration of L-glutamine applied to the olfactory epithelium (horizontal bars) increases from 10-7 M to 10-3 M, a concomitant increase in activity is seen in the nerve. Upper traces show the integrated responses and lower traces show the raw (nerve) signal. (B) Semi-logarithmic plot of normalized data fitted to a three-parameter Hill equation (R2 = 0.99). For this example, the calculated EC50 = 3.11 µM, and the Hill co-efficient = 0.565). Please click here to view a larger version of this figure.
