Curcumin labels Aβ plaques within a minute. When we stained 5xFAD tissue with Cur, we found that Cur label Aβ plaques within 1 min. Although increased incubation time with Cur slightly increased the fluorescence intensity of Aβ plaques, the number of observed Aβ plaques was not significantly different between 1 min and 5 min staining time (Figure 1).
Cur can label Aβ plaques in cryostat-prepared, paraffin-embedded mouse and human AD tissue that colocalizes with Aβ-specific antibodies in mouse AD brain tissue. When we stained cryosection, paraffin-embedded sections (Figure 2A) and human AD tissue (Figure 2B), we observed Cur-labeled Aβ plaques in all types of tissue sections. Additionally, to confirm that Cur is binding to Aβ plaques, we first labeled the plaques with 6E10, followed by Cur staining. We observed that the Cur was completely co-localized with Aβ at the same plaques that bound the 6E10 (Figure 2C).
Cur labeled Aβ oligomers and intracellular Aβ aggregates. To check whether Cur could label Aβ oligomers, 5xFAD mouse sections were stained with an Aβ oligomer-specific antibody (A11), followed by Cur staining. We observed that Cur colocalized with A11 in Aβ plaques (Figure 3A). Similarly, Cur also colocalized with the 6E10 antibody in intracellular spaces (Figure 3B), indicating that it can label the intracellular Aβ.
Cur labeled Aβ more prominently than classical amyloid binding dyes. Aβ labeling by Cur was compared with commercially available amyloid binding dyes Thio-S, CR, FJC. The Aβ-specific antibody 6E10 was used as standard control. We observed that Cur labeled Aβ more prominently than the conventional amyloid binding dyes (Figure 4).
Cur derivatives bis-demethoxycurcumin (BDMC) and demethoxycurcumin (DMC), also present in turmeric extract, label Aβ plaques comparatively to Cur in 5xFAD brain tissue. Two other major components, such as BDMC and DMC, are present in turmeric extract. We tested whether these two compounds also label Aβ plaques similar to Cur. When 5xFAD mouse brain sections were stained with these derivatives, both BDMC and DMC also labeled Aβ plaques, parallelling Cur (Figure 5A). Different mounting media were investigated to check for interference with Aβ-imaging after staining with Cur. The fluorescent signal was intact in both aqueous and organic mounting media, such as DPX (Figure 5B). The staining of Aβ plaques with Cur was appropriate after immunofluorescent labeling and followed by counterstaining with Hoechst 33342 solution (1 mg/ml) or 4′,6-diamidino-2-phenylindole (DAPI, 1µg/ml). The immunofluorescent signals and counterstaining intensity were maintained after Cur staining (Figure 5C).
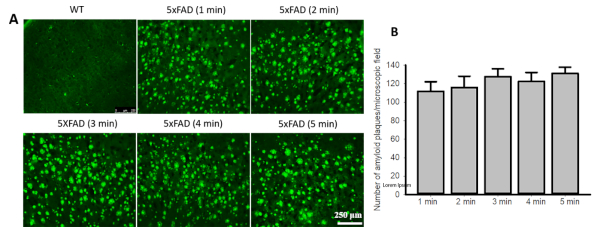
Figure 1: Curcumin labels Aβ plaques within a minute. (A). Brain sections from 5xFAD mice or human control and AD cortical tissue were stained with Cur (10 µM) for 1−5 min and the number of visible plaques were counted. (B). No difference was observed in the number of Aβ-plaque counts between the 1 min and 5 min staining times. Data are represented as mean ± standard error of mean (SEM). Please click here to view a larger version of this figure.
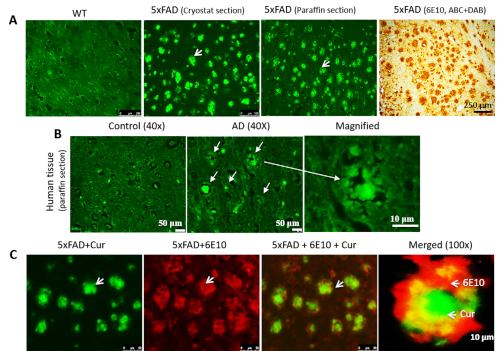
Figure 2: Colocalization of Cur with Aβ antibody in Aβ plaques. Cur can label Aβ plaques in cryostat-prepared, paraffin-embedded sections (A) and human AD tissue (B). The Aβ labeling paralleled the labeling with Aβ-specific antibody (6E10, DAB staining). (C). The 5xFAD sections were first labeled with Aβ antibody (6E10), followed by staining with Cur. Red = 6E10 bound by a secondary antibody tagged with Alexa fluorophore 594. Green = Cur. Cur completely colocalized with Aβ at the same plaques that bound to 6E10. Arrows indicate Aβ plaques. Scale bar indicates 50 µm (total magnification = 400x) in all three images on the left, and 10 µm (total magnification = 400x). Please click here to view a larger version of this figure.
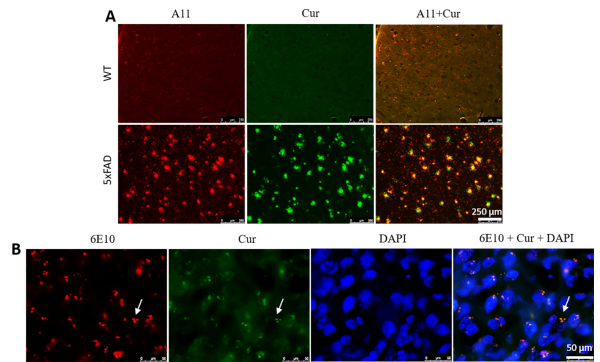
Figure 3: Curcumin labels Aβ oligomers and intracellular Aβ. (A) Aβ-oligomer was detected immunohistochemically using an Aβ-specific antibody (A11), followed by staining with Cur. Cur completely colocalized with Aβ, at the same oligomer where A11 binds. Scale bar = 250 µm and total magnification = 100x. (B) Similarly, intracellular Aβ was detected immunohistochemically using an Aβ-specific antibody (6E10), followed by staining with Cur. Note that Cur completely colocalized with Aβ in the same areas bound to 6E10. Scale bar = 50 µm and total magnification = 1,000x. Please click here to view a larger version of this figure.

Figure 4: Comparison of different amyloid binding dyes with Cur to label Aβ plaques. Cryostat sections (40 µm) from the cortex of 12-month-old 5xFAD mice were stained with Cur, Thioflavin-S, Congo red, Fluoro-jade C, and with 6E10 antibody. Cur labeled Aβ plaques more prominently than Thio-S, CR, and FJC. Arrows indicate Aβ plaques. Please click here to view a larger version of this figure.
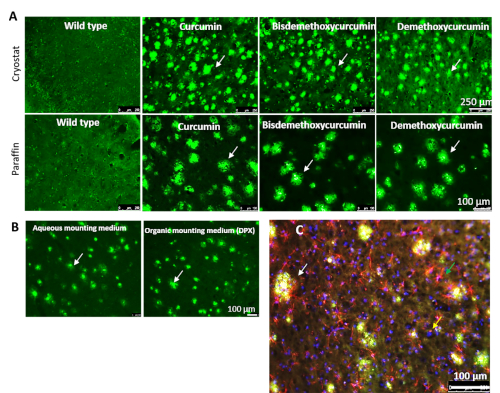
Figure 5: Cur-derivatives bis-demethoxy curcumin and demethoxy curcumin also label Aβ similarly to Cur. (A) Both cryostat and paraffin-embedded 5xFAD sections were stained with Cur-derivatives bis-demethoxy curcumin and demethoxy curcumin. Both these derivatives label Aβ plaques in a manner similar to Cur. (B) Both aqueous and organic mounting media (DPX) were used to mount tissue sections after labeling Aβ plaques with Cur. (C) Immunolabeled sections were used for Aβ labeling with Cur. The white arrows indicate Aβ plaques, the green arrow indicates activated astrocyte (GFAP), and the yellow arrow indicates nuclear stain (DAPI). Scale bars = 100 µm. Please click here to view a larger version of this figure.
| Features | Aβ-antibody | Curcumin | Thio-S | Congo red | Fluoro-Jade C |
| Duration of staining | ~24-48 h | ~1-5 min | ~10 min | ~60 min | ~30 min |
| Accessory chemicals | Secondary antibodies, several chemicals to make buffer, normal goat serum | Methanol | Ethanol | NaOH and ethanol | NaOH and ethanol, potassium permanganate |
| Cost | Costly: one Aβ-specific antibody vial requires ~$200-300 | Cost effective: ~$5-10/1 g Cur and can be applied for many tissues | Cost effective: ~$5/1 g, can be applied for few tissues | Cost effective: >$5/1 g Congo red, and can be applied for several tissues | Costly: ~$15.500/1 g FJC powder |
| Specificity | Different antibodies are required for Aβ oligomers and fibrils | Curcumin binds with Aβ oligomers and fibrils | Can bind only fibrils, not monomers, or oligomers | Can only bind with Aβ-protofibrils and fibrils16,17 | Can only bind with Aβ-fibrils and degenerated neurons |
| Stability | Depending on the dye attached to the secondary antibody | Very stable, even at room temperature when bound with Aβ | Stable in methanol | Stable in ethanol | Not stable |
| Care after staining | Needs extra care after staining, such as being kept in the dark and frozen all the time | Not as light sensitive and more stable at room temperature | Light sensitive | Not light sensitive | Light sensitive |
| Microscope required | Compound light or fluorescent (depending on use of secondary antibody) | Fluorescent | Fluorescent | Light microscope or polarized microscope or polarize filter | Fluorescent |
| Background staining | Generally, no background | Very low background | High background due to binding with lipid membrane or lipid compounds in cell | Low background | High background |
| In vivo Aβ-imaging | May not be applicable | Highly applicable | May not be applicable | May not be applicable | May not be applicable |
Table 1: Comparison of Aβ labeling with different amyloid binding dyes and Cur7.
