The described methodology has been used to study long-lasting forms of LTP/LTD and its associative interactions such as synaptic tagging and cross-capture from acute hippocampal slices of adult rats.23 This technique has proven effective for experiments with both rats (Wistar) and a variety of mouse strains30,31. The methodology has been used successfully for stable LTP recordings of up to 8-12 hr.32
The ‘tag’ set by the weak tetanization of one input (S1) captures the ‘PRPs’ induced by the strong tetanization of another independent but overlapping input (S2; Figure 3B, filled circles) thereby transforming the otherwise decaying form of LTP (early-LTP) in S1 into a long-lasting one (Figure 3B, open circles) (For comparison of early-LTP induced by WTET see20,33). The PRPs captured by the weak tetanization set tag need not necessarily come from the STET-induced late-LTP but can also be provided by the SLFS-induced late-LTD. This type of positive associative interaction between LTP and LTD is referred to as ‘cross-tagging/capture’. The WTET-induced early-LTP in S1 gets reinforced to late-LTP (Figure 3C, open circles) by capturing the PRPs provided by the SLFS-induced late-LTD in S2 (Figure 3C, filled circles). Statistically significant potentiation or depression was maintained in S1 and S2 in both cases when compared to its own baseline (Wilcoxon test; P < 0.05).
For the tag-PRP interaction to occur, the temporal order of the two events (weak-before-strong/strong-before-weak) is not crucial as long as the time window between the two events remains within the range of 30-60 minutes. It would be wise to include a third, independent but overlapping synaptic input and use it as a baseline control to monitor the stability of recordings. The electrical stimulation protocols used to induce early- and late forms of LTP/LTD must be validated in single-input experiments for consistency and reliability before using them in STC experiments. We would also like to emphasize the importance of slice preparation methodology described in the protocol since the success of these experiments relies heavily on the quality of the slices.
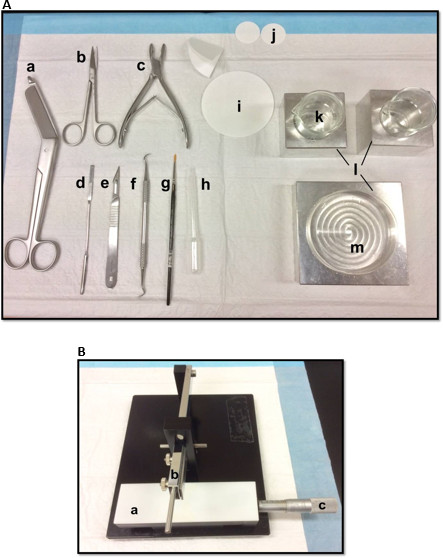
Figure 1. (A) Tools used in the dissection of hippocampus: (a) Bandage Scissors (b) Iris scissors (c) Bone rongeur (d) Thin spatula, (e) Scalpel number 11 (f) Sickle scaler (g) Soft-bristle paint brush (h) Plastic Pasteur pipette (i) filter paper (85mm) (j) filter paper (30mm) (k) Glass beakers (l) Aluminum cooling blocks to fit petri dish and beakers (m) Petri dish. (B) Manual tissue chopper. (a) Platform (b) Cutting arm with blade-holder (c) Vernier micrometer, resolution 10 microns. Please click here to view a larger version of this figure.

Figure 2. Electrophysiology set-up for field-potential recordings consisting of (A) stimulators (b) a differential amplifier (c) an analog-to-digital converter (d) Oscilloscope (e) computer with acquisition software (f) Vibration-resistant table-top (g) microscope with >4x magnification (h) interface brain-slice chamber (i) a perfusion system for ACSF and carbogen supply (j) temperature controller (k) an illumination source (l) manipulators with electrode holders. (B) Interface brain-slice chamber. (C) & (D) Hippocampal slices in the interface chamber. (E) Stainless steel electrode sealed in a glass capillary. Please click here to view a larger version of this figure.
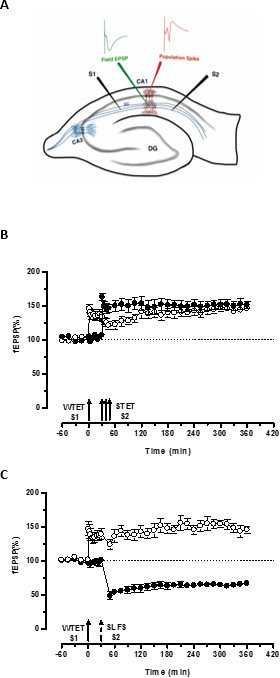
Figure 3. (A) Schematic representation of a transverse hippocampal slice and electrode location for field-potential recording: In this representation, two stimulating electrodes (S1 and S2) are positioned in the stratum radiatum of the CA1 region to stimulate two independent but overlapping synaptic inputs onto CA1 pyramidal neurons. Two extracellular recording electrodes, one to record field-EPSP (excitatory post-synaptic potential) from the apical dendritic compartment and another to record somatic population spike from the pyramidal cell bodies, are located in the stratum radiatum and stratum pyramidale respectively. CA1- cornu ammonis region 1, CA3- cornu ammonis region 3, DG- dentate gyrus, SC- Schaffer collateral fibers, S1- stimulating electrode 1, S2-stimulating electrode 2. (B) Weak before strong paradigm to study STC: Weak tetanization (WTET) is applied to S1 (open circles) for inducing early-LTP followed by strong tetanization (STET) of S2 (filled circles) at 30 min to induce late-LTP. The early-LTP in S1 gets reinforced to late-LTP showing tagging and capture interaction (n = 6). (C) Weak before strong paradigm to study cross-tagging: Early-LTP is induced by WTET in S1 (open circles) followed by the induction of late-LTD in S2 (filled circles) using SLFS after 30 min. In S1, the early-LTP is transformed to late-LTP lasting 6 hr showing cross-tagging and capture (n = 6). Single arrow represents weak tetanization applied for inducing early-LTP. Triplet of arrows represents strong tetanization for inducing late-LTP. The broken arrow represents the time point at which SLFS was applied to the representative synaptic input to induce late-LTD. Error bars indicate SEM. Please click here to view a larger version of this figure.
