The protocol described above is an efficient means to isolate CFTR-enriched microsomes, with almost complete recovery of CFTR during the cell breakage and preparation of the crude microsomes (Figure 1). Other cell breakage methods may also be employed effectively. We have utilized a French pressure cell, and other high-pressure/cavitation devices (also in combination with impacting against a ruby target) with equal efficiency. For convenience and low initial cost of the equipment, we find the bead-beating method the best.
Using LPG to solubilize and purify CFTR yielded 80 μg protein/L culture at >90% purity (Figure 2). The high yield was due to efficient solubilization of CFTR by LPG (compare Figure 2b, lanes 2 and 4). In addition, efficient and tight binding to the column resulted in minimal loss of CFTR in the unbound fraction and the absence of CFTR in the wash fractions (Figure 2, lanes 3, 5, and 6). The eluted protein had a purity of >90%, estimated by Coomassie-stained SDS-PAGE gels and using densitometry of the CFTR and contaminant bands. Gel permeation chromatography (GPC) separated LPG-purified CFTR from low-molecular weight contaminants (Figure 4, lower panel).
The protocol for CFTR purification using DDM gives purity of about 60% and yield of roughly 50 μg/L (Figure 3). Electron microscopy (EM) of negatively stained fractions from the GPC eluting at about 10 ml (Figure 4) showed that DDM-purified CFTR contains aggregates of 20-30 nm diameter as well as smaller particles of 10 nm diameter (data not shown). It is possible that the small aggregates can reversibly associate and dissociate as ultrafiltration with a 1 MDa cut-off filter failed to remove the EM-detectable aggregates. LPG-purified material did not adsorb to a glow-discharged grid, hence was studied by cryo-EM of unstained fractions. This showed a very homogeneous particle population of a relatively small size (6-8 nm diameter, data not shown).
Finally, the ATPase activity of the purified proteins was measured (Figure 5). As a member of the ABC protein family, CFTR has two nucleotide-binding domains (NBDs) capable of binding and/or hydrolyzing ATP. The data indicate that the purified protein was not able to hydrolyze ATP in the LPG-solubilized state and showed weak ATPase activity in the presence of DDM (Figure 5, unfilled bars). After the addition of lipids, and detergent removal, ATPase activity was 4-fold higher for samples that had been purified in DDM (13 nmol ATP/min/mg protein). The addition of lipids and removal of LPG similarly restored activity to CFTR that had been isolated using LPG, but with a final lower rate (1.5 nmol ATP/min/mg protein) than the DDM-purified and reconstituted material.

Figure 1. Monitoring levels of chicken CFTR in cell lysate (CL), supernatants (S) and pellets (P) during various centrifugation steps used for microsome isolation and washing. SDS-PAGE gels were visualized using the in-gel fluorescence of the GFP tag. The supernatant after cell breakage and centrifugation at 14,000 x g contains virtually all the CFTR (including degradation products). Ultracentrifugation at 200,000 x g sediments all the full-length CFTR leaving some fragments in the supernatant. Ultracentrifugation at 100,000 x g of salt-washed microsomes pellets nearly all the CFTR with the removal of some further fragments.
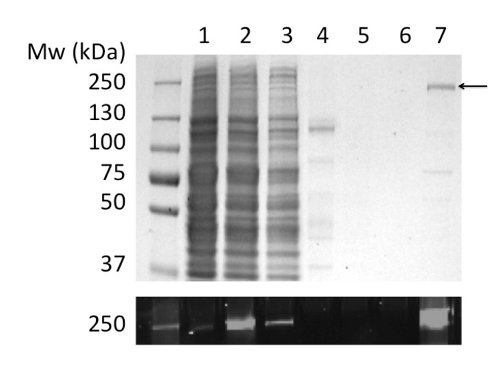
Figure 2. Purification of chicken CFTR in LPG by immobilized metal ion affinity chromatography. Fractions were analyzed by SDS-PAGE followed by Coomassie staining (upper panel) and fluorescence detection of the GFP tag (lower panel). Tracks: (1) Microsomes. (2) LPG-solubilized microsomes. (3) Unbound material. (4) Insoluble material. (5) & (6) 40 and 100 mM imidazole washes. (7) Material eluted with 400 mM imidazole.
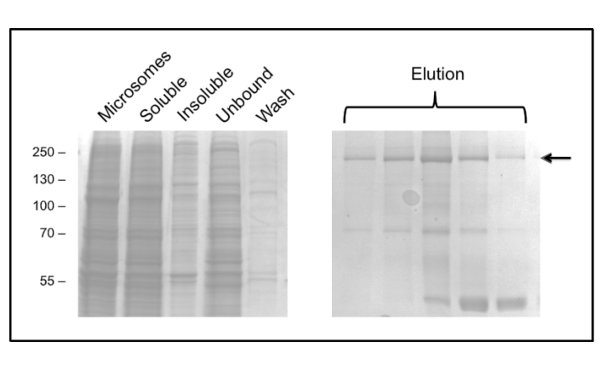
Figure 3. Purification of chicken CFTR in DDM by immobilized metal ion affinity chromatography. Fractions were analyzed by SDS-PAGE followed by Coomassie staining. The left hand panel shows fractions prior to elution. Several consecutive elution fractions are shown in the right hand panel with CFTR indicated by the arrow. Later fractions are enriched in a 40 kDa contaminant, which has been identified by mass spectrometry as ribosomal protein L3.
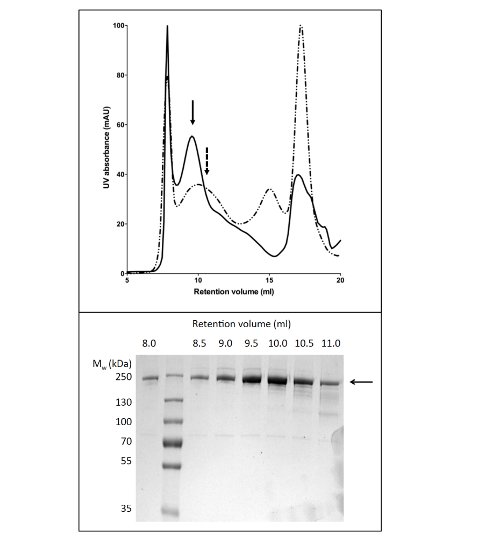
Figure 4. Purification of chicken CFTR by gel permeation chromatography. CFTR purified by Ni-affinity chromatography was concentrated and applied to a GPC column. The elution profile for CFTR (upper panel) purified in buffer containing LPG-14 (solid line) or DDM (dashed line) are overlaid. SDS-PAGE (lower panel) revealed that CFTR eluted between 8 and 11 ml.
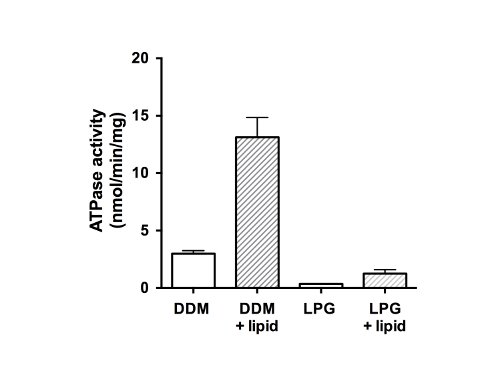
Figure 5. ATPase activity of purified chicken CFTR fractions. Protein purified in DDM or LPG was assayed using a modified Chifflet assay26 in the presence of a cocktail of ATPase inhibitors to eliminate any background ATPase activity from F-, P- and V-type ATPases (unfilled bars). The rate of ATP hydrolysis was also measured after detergent removal and lipid addition (filled bars). The plot shows the mean and standard deviation (n=3). Differences between mean values for ATPase activity in presence and absence of lipid, and difference between activity in DDM and LPG are significant to p<0.05.
