The assay consists of monitoring pyranine fluorescence as a function of time while a proton gradient is generated. To that purpose, samples are subjected alternatively to illumination (hence proton pumping by the BR is triggered) and then to fluorescence measurement using the excitation and emission wavelengths of pyranine (λex = 455 nm and λem = 509 nm).
Figure 1 shows a representative control obtained with the assay, in the absence of MexA/MexB, verifying that the proton gradient generated by the BR is reversible. After one cycle of acidification, proteoliposomes are kept in the dark for 45 min in order for the BR to stop pumping protons (protons diffuse slowly and passively across lipid bilayers following their concentration gradient). After this recovery time, activation of BR is possible, simply by illuminating the same suspension again.
Figure 2 corresponds to an actual transport measurement. A negative control with protein free liposomes shows that, as expected, the pH is constant upon illumination (Figures 2A and 2B, orange circles). However, proteoliposomes containing BR in their membranes do pump proton, thus the pH inside the liposome decreases and a stable gradient is built (Figures 2A and 2B, purple squares). Grey triangles and red diamonds (Figure 2A) represent pH inside of proteoliposomes containing BR and MexB, in the presence or in the absence of Hoechst 33342, respectively. We observe a substrate-independent activity where the proton gradient is only partially dissipated by MexB counter-transport. We attribute this observation to a basal activity of MexB.
In order to test the effect of MexA on the activity of MexB, MexA was added to the reconstitution, first in the absence of any substrate (Figure 2B, blue diamonds). Again, the proton gradient generated by the BR is only partially dissipated. The actual transport measurement is realized on the very same sample. Beforehand, the proton gradient must be reinitialized and the substrate must be added in the suspension in order to reach its binding site in the protein. To that purpose the suspension is incubated in the dark in the presence of the substrate for 45 min. Upon subsequent illumination, one can see that the proton gradient generated by the BR is now totally dissipated by MexAB (Figure 2B, green triangles), as a consequence of substrate transport by the pump.
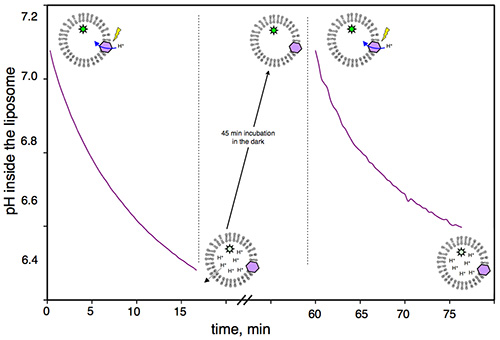
Figure 1. Reversibility of the proton gradient built by BR illumination: Pyranine fluorescence of BR proteoliposomes measured as a function of time. From 0-15 min, BR pumps protons as a result of light activation. From 15-60 min, proteoliposomes are incubated in the dark; during this time, protons passively move out of the liposomes until the intravesicular pH and extravesicular pH are equal. From 60-75 min, BR is still functional and pumps protons upon illumination.
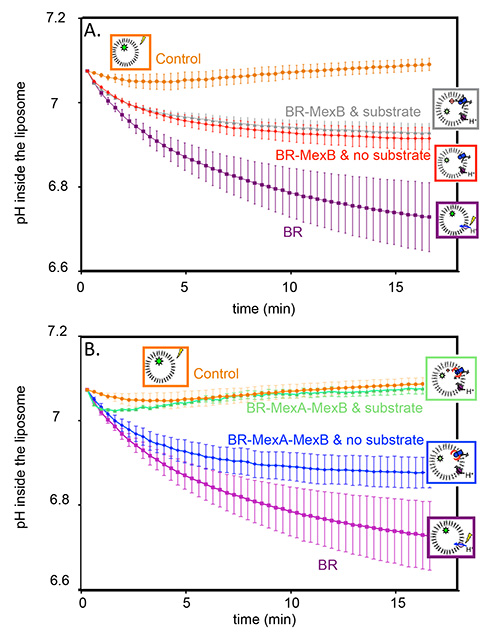
Figure 2. Transport assay: pyranine fluorescence, converted to the corresponding pH variations, as a function of time. A) pH inside of control liposomes (orange circles); pH inside of proteoliposomes containing BR in the membrane (purple squares); pH inside of proteoliposomes containing BR, and MexB in the membrane without Hoechst 33342 (red diamonds); pH inside of proteoliposomes containing BR and MexB in the membrane with Hoechst 33342 (grey triangles). B) pH inside of control liposomes (orange circles); pH inside of proteoliposomes containing BR in the membrane (purple squares); pH inside of proteoliposomes containing BR, MexB and MexA in the membrane without Hoechst 33342 (blue diamonds); pH inside of proteoliposomes containing BR, MexB and MexA in the membrane with Hoechst 33342 (green triangles).
