All zebrafish care protocols and experiments were performed under Institutional Animal Care and Use Committee (IACUC) protocol A2009-11-01.
1. Morpholino and Larval Injection Dishes
Experimental duration: * (10-15 minutes)
Degree of difficulty: *
- For egg injections, prepare a 2% agarose solution in sterile water and microwave. When the solution has cooled pour some of it into an extra deep Petri dish (Fisher Scientific) until it is half full. Cool on ice and make sure the plate is level.
- Once the dish has cooled, pour a 15 mL top layer of 2% agarose. Spray an egg injection grooved plastic mold (Adaptive Science Tools) with sterile water from a spray bottle. Carefully place the mold groove side down into the hot agarose. When the agarose has cooled, use a metal flat spatula (VWR Scientific) to dissociate the mold from the agarose. Slowly remove the grid from the agarose. Wrap embryo injection dishes in Parafilm (VWR Scientific) and store inverted at 4 °C.
- For fish larval injections, prepare a 2% agarose solution as described. Pour the solution into a standard size Petri dish (VWR Scientific) and set aside until it solidifies. Wrap larval injection dishes in Parafilm and store inverted at 4 °C.
2. Fungal Culture Preparation
Experimental duration: **(30 minutes)
Degree of difficulty: **
- Prepare yeast extract-peptone-dextrose (YPD) agar plates: 10 g/liter yeast extract, 20 g/liter peptone, 20 g/liter dextrose, and 20 g/liter agar into and autoclave. For YPD liquid prepare 10 g/liter yeast extract, 20 g/liter peptone, 20 g/liter dextrose and autoclave 20-30 minutes at 121 °C.
- Two days prior to fish infections prepare a streak plate from frozen stocks of Candida albicans cultures onto YPD agar to obtain single colonies.
- Incubate at 37 °C overnight.
- Put 5 mL YPD broth into 16 x 150 mm culture tubes (VWR Scientific).
- The day before the fish infections pick 1 small colony on the YPD agar with a wooden dowel (VWR Scientific). Put the stick in the culture tube and swirl around to re-suspend colony in the YPD liquid.
- Grow overnight at 37 °C on a TC-7 Tissue Culture Roller Drum equipped with a 14-inch test tube wheel (New Brunswick Scientific).
- The next day, spin down 1 mL of culture at 14000x g for 1 minute in a 1.7 mL tube (Axygen). Remove the supernatant and resuspend the pellet in residual liquid by vortexing (VWR Scientific).
- Add 1 mL 1x Phosphate Buffered Saline (VWR Scientific) and spin as previously described.
- Repeat the PBS wash 3 times.
- Dilute 1:100 dilution in 1x PBS (10 μL of washed C. albicans into 990 μL 1x PBS).
- Count on a Hemocytometer (VWR Scientific).
- Dilute to 107 cells/mL.
3. Zebrafish Infections
Experimental duration: **** (1-3 hours)
Degree of difficulty: ****
- Collect embryos according to section 5 and store in egg-water, 60 mg/L instant ocean salts (Fisher Scientific) in sterile deionized water.
- Using a dissecting microscope such as the Olympus SZ61 (Olympus), dechorionate embryos on infection day. Use Dumont Dumoxel tweezers (VWR Scientific) to pull the chorion27 apart like opening a bag of chips, or gently poke the tweezers in the closed position into the chorion and then slowly open them. The fish should pop right out of the chorion.
- Swirl the extra deep Petri dish with the lid on to move fish into the center of the dish. Transfer fish to the lid of the dish. Remove egg-water and replace with fresh media. Add fish to media.
- Warm larval injection dishes in an incubator at 28 °C. Prepare tricaine methane sulfonate (Western Chemical Inc.) dilution at 200 μg/mL for anesthetizing fish.
- Count out the desired number of larvae for infection (20-50 fish). Put fish in tricaine methane sulfonate solution and wait 1-2 minutes until they stop moving.
- Turn on the MPPI-3 injection unit (Applied Scientific Instruments). Make sure the pressure switch is on “pulse” and the “pulse duration” is set to 9 with 3 PSI for the backpressure unit. Open the valve to the nitrogen tank until the pressure on the injection unit reads 30 PSI.
- Load a pulled micropipette28 with 5 μL of vortexed Candida albicans at a concentration of 1×107 cells/mL. Place the micropipette in the micropipette holder (Applied Scientific Instruments). Fill an empty extra deep Petri dish with water. Move the micropipette until the tip is within view just touching the surface of the water via dissecting microscope and use Dumont Dumoxel tweezers (VWR Scientific) to clip the needle about 3mm from the tip of the pulled pipette.
- Press the foot switch (Applied Scientific Instruments) to verify the needle has been clipped. You should see the liquid disperse if you were successful. The diameter of the liquid bolus should be no bigger than the diameter of the pupil of the prim25 zebrafish larva (0.21 mm, yielding a sphere of 4.9 nL in volume). Adjust the pressure accordingly if the liquid bolus is too big or too small.
- Anesthetize the fish in tricaine methane sulfonate (200 μg/mL). Once the fish have stopped moving, swirl the dish with the lid on until the fish are in the center. Collect 50 fish using a transfer pipette (Fisher Scientific). Tap the side of the pipette to settle the fish towards the tip. Gently pipette fish onto the larval agarose dish using as little liquid as possible.
- Line the fish up with a smooth glass rod (be careful not to crush them!). Aspirate as much liquid as possible off of the dish. Use a KimWipe to wick away any moisture.
- Position the larval agarose dish with fish under the microscope until you get a good view of the fish and the needle at the same time. Move the glass needle towards the fish. Zoom in on both the fish and needle as you position the needle towards the fish. Carefully move the glass needle into the otic vesicle27, 29 (ear) of the first fish. You’ll know once you’ve moved the needle into the fish because when you push the foot switch (Applied Scientific Instruments), the hindbrain ventricle will lift up slightly.
- Press the foot switch (Applied Scientific Instruments) and watch the liquid disperse inside the fish’s hindbrain ventricle27. Retract the needle from the fish and move to the next fish. Repeat procedure until you have injected all of the fish on the plate. Remove any dead fish and inject replacements to keep the number correct (50 fish). For each group of 50 fish injected a new microinjection needle must be prepared, or the C. albicans settles in the bottom of the needle and clogs it, or throws off the concentration injected.
- Wash the fish off of the dish by angling the dish and spraying egg-water onto the fish into a clean extra deep Petri dish containing 60 mL egg-water. It is important not to leave the fish on the agarose for longer than 15-20 minutes or they will dry out and die.
- Repeat entire injection process until finished. Don’t forget PBS and non-pathogenic injected controls.
- When finished, keep the fish at 28 °C.
- Close the nitrogen tank valve. Move the injection unit switch from “pulse” to “continuous” to relieve pressure in the tank line, the pressure should drop to zero at this point. Switch injection unit back to “pulse”. Turn off the injection unit and clean up the work area.
4. Preparing the Fish for Imaging
Experimental duration: ** (30 minutes)
Degree of difficulty: **
- Prepare tricaine methane sulfonate solution as before (200 μg/mL). Get out infected fish, and transfer them into tricaine.
- Prepare 47.9 mL 0.4% low-melt agarose (VWR scientific) in egg-water. Heat the solution by microwaving. Cool to 37 °C and add tricaine methane sulfonate (200 μg/mL) to mixture
- Once fish are immobilized in tricaine methane sulfonate, pipette individual fish into a Petri dish (VWR scientific) containing 0.4% low melt agarose (VWR Scientific).
- Next, move the fish from low melt agarose into individual wells of a glass bottom dish (MatTek Corporation) for imaging. Use as little low melt agarose as possible so the fish lies flat on the bottom of the dish. Only use enough of the tricaine-agarose to fill the inner circles of the glass bottom dish.
5. Modifications Related to JoVE Protocols
Micropipettes for Microinjection
Experimental duration: * (10-15 minutes)
Degree of difficulty: *
- Pull hollow glass rods BF120-69-10 (Sutter Instruments) using a Flaming Brown Micropipette Puller Model P-97 (Sutter Instruments) according to Yuan et al.30. Choose program #7 with the following conditions Heat = 470, Velocity = 120, Time = 200. The resulting needle is 8 mm from bevel to tip and, once clipped at 3 mm above the tip it has a diameter of approximately 10 μm.
- Load a glass micropipette into the brackets. Select “Pull.” The heating filament will heat the needle according to the program parameters and the glass rod will separate into two pulled micropipettes. Store micropipettes in a pipette storage box (Sutter instruments).
Embryo Collection, Morpholino Injection and Maintenance
Experimental duration: *** (1-2 hours)
Degree of difficulty: ***
- Collect embryos according to the methods of Rosen et al.31. Use a plastic sieve (Wares of Knutsford) to collect eggs from the spawning tank (Aquatic Habitats). Hold the sieve upside down over an extra deep Petri dish and rinse with tank water to collect eggs.
- For morpholino preparation, add 300 μL sterile water to 300 nanomoles of morpholino (GeneTools, LLC). This gives a working stock of 1.0 mM.
- Verify the concentration of the morpholino by using a Nanodrop (Thermo Scientific). Select “nucleic acid and change the sample type to “other” and wavelength to 265. Enter the constant by multiplying the molecular weight of the morpholino by 1000 divided by the absorptivity coefficient listed in the morpholino oligo property sheet.
- Blank the nanodrop with 2 μL 0.1 N HCl. Dilute 5 μL of the morpholino solution into 95 μL of 0.1 N HCl (20x dilution). Place 2 μL of the dilution on the nanodrop pedestal and click measure. Multiply the concentration by the dilution factor (20). This gives a working concentration of morpholino. To calculate the millimolar concentration divide the concentration obtained by the molecular weight of the morpholino.
- Prepare a working stock of morpholino in Danieau buffer (58 mM NaCl, 0.7 mM KCl, 0.4 mM MgSO4, 0.6 mM Ca(NO3)2, 5.0 mM HEPES, pH 7.6) and 0.01% phenol red (VWR Scientific).
- For morpholino injection work follow the protocol according to Rosen et al.31 and Yuan et al.30. Rinse morpholino injection dishes with egg-water and warm at 28 °C for 15 minutes. Remove water from injection dishes and line-up one-cell stage embryos into the grooves of the injection dish (prepared from Section 1) with a transfer pipette. Each plate will hold approximately 250 eggs. Inject 1-2 cell stage embryos with morpholino. Eggs will remain in the one-cell stage for 15 minutes27.
- Rinse injected eggs with egg-water when finished (sterile deionized water with 60 mg/L Instant Ocean salts) and place in 60 mL egg-water in extra deep Petri dishes. Morpholino injection dishes can be reused. Rinse with egg-water when finished and store inverted at 4 °C.
- For storage of embryos add 60 mL egg-water to dish. Count out 110 embryos with a transfer pipette into separate extra deep Petri dishes with 60 mL egg-water. Add 0.00003% methylene blue to prevent microbial growth and incubate embryos at 28 °C.
- To speed up development of embryos keep embryo dishes at 33 °C for 24 hours and stage according to Kimmel et al.27, allowing embryos to develop to a head trunk angle of 75° (Prim 25 stage27).
- For larval infection work keep embryos at 28 °C after injecting with C. albicans.
- Each day replace embryo dishes with 60 mL fresh egg-water.
Imaging
Experimental duration: *****(1-5 hours)
Degree of difficulty: ***
- Prepare fish in low melt agarose (according to section 4.2) with tricaine in glass bottom imaging dishes (MatTek Corporation) as described earlier. Place the dish on an Olympus IX-81 (Olympus) inverted microscope stage. Bring the fish into focus under 4x magnification under differential interference contrast (DIC) light. Images can be captured at 4x, 20x, and 40x magnification in DIC, TRITC (tetramethyl rhodamine isothiocyanate), and FITC (fluorescein isothiocyanate) filter settings.
- Use an Olympus IX-81 inverted microscope with an FV-1000 laser scanning confocal system according to Ariga et al.32. Conduct time courses at 40x magnification by setting Z-stacks with 1-1.5 μm slices every hour of infection.
- For long time course imaging of 2 hours or more, place a layer of low melt agarose on top of the fish in the imaging dish every 2-3 hours. This prevents the agarose from drying out and crushing the fish while it is being imaged. For lengthy time courses with a small number of fish, use a heated stage (Bioptechs Inc.) to maintain 28 °C during infections.
6. Representative Results
An example of a successful hindbrain ventricle C. albicans infection in a zebrafish larva at 5 hours post-infection (hpi) and 24 hpi is shown in (Figure 1). Macrophage-like cells with engulfed C. albicans are seen in the hindbrain ventricle at 5 hpi. By 24 hpi, C. albicans is inside macrophage-like cells in the dorsal tail tissue indicative of disseminated candidiasis. This infection result is highly dependent upon an accurate injection of 10-15 yeast-form C. albicans into the hindbrain ventricle. Screening of infected fish immediately post-injection can ensure this.
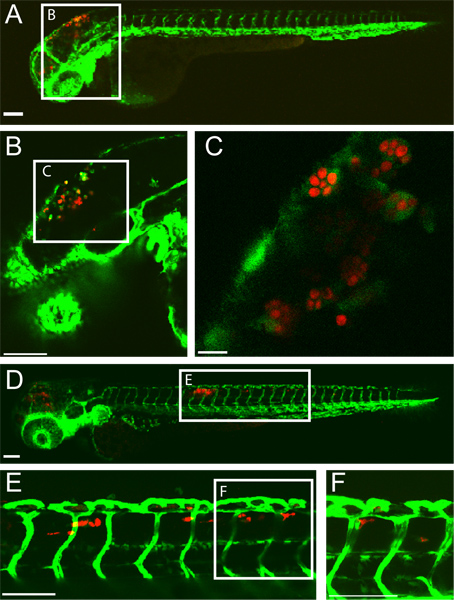
Figure 1. Transgenic fli1:EGFP22, 33 larva infected with CAF2-yCherry Candida albicans and imaged intravitally by confocal microscopy. (A-C) 5 hours post-infection (A) Infected larva with EGFP-expressing macrophage-like cells at the site of infection (hindbrain ventricle) Scale bar =100 μm. (B and C) Higher magnification images of same fish, showing C. albicans within phagocytes. Scale bar = 100 μm for B and 10 μm for C. (D-F) 24 hours post-infection (D) Infected larva with disseminated candidiasis with CAF2-yCherry C. albicans inside EGFP macrophage-like cells in the dorsal tail tissue. Scale bar = 100 μm. (E and F) Higher magnification images of same fish, showing C. albicans in tail tissue. Scale bar = 100 μm.
