Growth assays, like ACT1-CUP1, require the visual, comparative assessment of multiple colonies. Here, each strain was grown to saturation overnight, diluted to an OD600 of 0.5, and plated on 20 plates containing a range of copper concentrations from 0 mM to 1.1 mM CuSO4 (Figure 3). This range is smaller than that listed in the protocol as it allowed for the full assessment of the impact of the QSPs and ACT1-CUP1 reporters used and described below. The plates were imaged and scored (Figure 3 and Supplemental Table 3).
For this representative experiment, the yeast background includes a disrupted ADE2 gene, resulting in the colonies being varying shades of red (Figure 3A,B). This common yeast disruption in the adenosine production pathway causes a buildup of a red-pigmented precursor for adenosine. Thus, the red color is an indicator of yeast colony maturity (i.e., the amount and age of the cells present). For the purposes of an ACT1-CUP1 assay, the red color can serve as an indicator of contaminating fungal species if white or yellow in color. The color can be a secondary confirmation of copper tolerance as more viable colonies will be a deeper red shade.
When plating with a pin replicator, there are several common aberrations. First, it is possible to create colonies that are oval-shaped if the replicator is lifted up while moving left or right (Figure 3B, orange arrow). In addition, microcolonies can also form from small droplets of cell solution if the pin replicator is brought in at an angle or if shaken above the plate (Figure 3B, red arrow). Often innocuous, occasionally, the microcolonies can mix with a stamped colony, and this prevents interpretation of that colony. Additional plating issues include insufficient wait time after sterilization for the replicator pins to cool and insufficient contact between the plate and the pins such that poor transfer of the culture media occurs. In both cases, few, if any, cells will grow on the plate, including for the control strains containing the wild-type reporter. As with any growth assay, regardless of the method of plating, it is important to perform the ACT1-CUP1 in triplicate to confirm the results are consistent and repeatable. Ideally, these different replicates should be performed on copper plates prepared at different times to ensure reproducibility.
For these representative experiments, the A3c reporter was selected to highlight the impact of a non-consensus sequence on copper tolerance in the absence of additional spliceosome perturbations. A3c significantly reduces the amount of CUP1 mRNA produced as it perturbs the ability of the spliceosome to recognize and utilize the 5' SS15. Yeast with the A3c reporter survived to 0.15 mM Cu2+, while the wild-type reporter cells maintained viability to the end of the range of copper concentrations tested (Figure 3C). The wild-type reporter containing cells grow to 2.5 mM Cu2+ without an impact on viability (data not shown).
U6 snRNA is an essential catalytic component of the spliceosome. Multiple studies have used ACT1-CUP1 to study the effect mutations can have on this RNA16,32,34. Duplicated for this study, three U6 snRNA sequences were studied, namely the wild-type sequence (WT), position 57 substituted from a uridine to a cytosine (U57c), and position 57 substituted from a uridine to an adenosine (U57a) (Figure 3 and Figure 4). Combined with ACT1-CUP1 reporters that impact the catalytic steps in splicing, it was determined that U57c favors the first catalytic step, and U57a favors the progression to the second step16,32.
To set up for the ACT1-CUP1 assay, a strain was created with the genomic copy of U6 snRNA deleted and the U6 snRNA wild-type or mutated sequences included on plasmids. As U6 snRNA is an essential component of the cell, three separate transformations were used to first knock-out the genomic U6 snRNA while maintaining cell viability, subsequently introduce mutated U6 snRNAs on plasmids, and, finally, add the ACT1-CUP1 reporters. For the first transformation, a cup1Δ yeast strain was used in the knock-out of the genomic copy of U6 snRNA while simultaneously adding WT U6 snRNA on a URA selection marker plasmid to maintain viability. A subsequent transformation added either wild-type or mutated U6 snRNA on a TRP selection marker plasmid, selecting against the URA marker via 5-FOA selection. Thus, three cup1Δ yeast strains were generated, each with one of the U6 snRNA sequences, namely WT, U57a, and U57c. A final yeast transformation for each strain added one of the three different ACT1-CUP1 reporter plasmids to be used in this experiment. The reporters selected were the wild-type reporter, A3c, and a mutation of the branch-site adenosine to guanine (BS-G). A total of nine strains were generated for this experiment, each containing one U6 snRNA sequence and one ACT1-CUP1 reporter (Supplemental Table 4 and Supplemental Table 5).
The results showed that the different bases at U6 snRNA position 57 have unique impacts on the spliceosome in combination with a 5' SS or BS mutation (Figure 4). Both the A3c and BS-G reporters primarily inhibit splicing by stabilizing the first catalytic step conformation14,15. Thus, U57c is an additive mutation decreasing copper tolerance in combination with either of these reporters (Figure 4B). In contrast, U57a increases copper tolerance because it promotes progression to the second step (Figure 4B)16,32. The BS-G reporter strain's decreased copper tolerance with U57a compared to U6 snRNA WT highlights a likely secondary impact of BS-G on the second step of splicing16.
These results also highlight the qualitative nature of this assay and why wild-type query and reporter sequences should be tested. While the general pattern of increased or decreased copper tolerance holds true for these U6 snRNA mutations compared to wild-type U6 snRNA, the exact concentration of copper to which the cells survive can differ between studies (Table 1) and did differ between these representative data and other published results. This is likely due to the background of the strain containing the CUP1 deletion but can also be due to the general health of the strains before plating (i.e., how soon after generation of the strain or restreak from cryo the assay was performed), differences in how the plates are prepared, and variability between different incubators. A similar variance was noted in Mayerle et al. for Prp8 query mutations in the literature43. Thus, comparing copper tolerance trends can be performed for different ACT1-CUP1 assays, but the numeric comparison of the copper concentrations should be done only within the same lab and, at times, with the same set of copper plates.
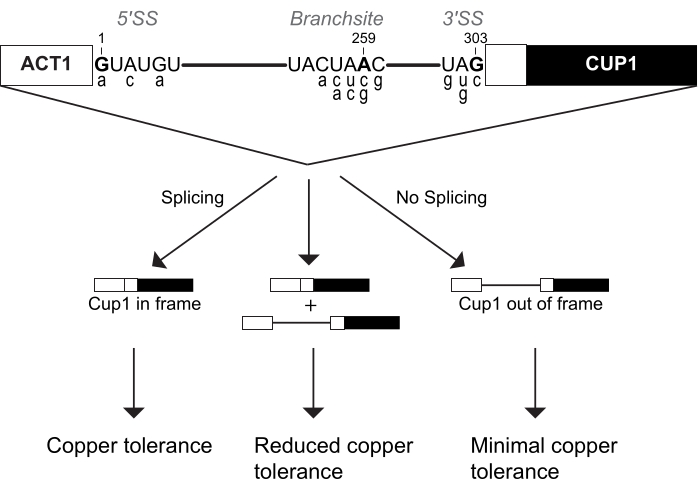
Figure 1: The ACT1-CUP1 reporter design. The concentration of spliced reporter directly correlates with yeast copper tolerance. The diagram of the reporter includes the three splice sites of the 5' splice site (5' SS), branch-site (BS), and 3' splice site (3' SS). The yeast consensus sequences for these sites are shown, with those targeted for cleavage indicated in bold and numbered based on their location in the intron. Commonly used non-consensus ACT1-CUP1 reporter sequences are shown in the lower case below their corresponding consensus sequence locations and are listed in Table 1. Please click here to view a larger version of this figure.
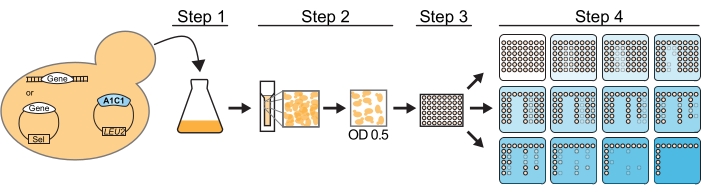
Figure 2: ACT1-CUP1 assay workflow. For this assay, a yeast strain must be cup1Δ leu2 and contain the desired ACT1-CUP1 reporter plasmid and the QSP gene, labeled Gene in the figure. The QSP gene must either be genomically inserted or on a plasmid. The protocol using a pin replicator involves four steps to prepare the yeast cells. Step 1 is to grow the cells to saturation. Step 2 is to measure the OD600 of the culture and dilute to an OD600 of 0.5. Step 3 is to distribute over a 96-well plate. Step 4 is to plate on plates containing increasing concentrations of copper (indicated with increasing blue color). Step 1, step 2, and step 4 would be identical for hand pipetting. Once plated, the plates are incubated for 3 days at 30 °C and then scored for viability. Please click here to view a larger version of this figure.

Figure 3: Representative views of ACT1-CUP1 data. For this experiment, yeast spliceosomes are challenged with two different U6 snRNA mutations and tested in the presence of three non-consensus reporters. Three replicates of this assay are presented on the same plates for demonstration purposes. It is recommended for this assay to do replicates on separate plates and on separate days. (A) A completed assay with a copper gradient of 0 mM to 1.1 mM over 20 plates is shown. As the copper concentration increases, strains with lower splicing show decreased confluency to the point where the copper concentration becomes lethal. The yeast background was ade2, and, thus, mature colonies are red in color. (B) Comparison of the same plates at 0 mM and 0.1 mM CuSO4 imaged with a handheld camera versus a digital imaging system. This is an example of how a number of non-consensus reporters can be compared side by side and with the wild-type reporter. Common observations on plates include a spurious drop of culture media that fell from the pin replicator (red arrow) and slight ovaling of the colonies due to sliding of the pin along the surface of the plate or movement of the plate before the drop of culture has dried sufficiently (orange arrow). (C) Example of the A3c reporter's effect on cell viability compared to the wild-type reporter. Images such as those shown in (B) are cropped and aligned to highlight the growth differences at different copper concentrations. The copper tolerance of the A3c reporter strain decreases to 0.15 mM Cu2+ compared to the wild-type reporter strain's viability to the end of the range tested. Please click here to view a larger version of this figure.
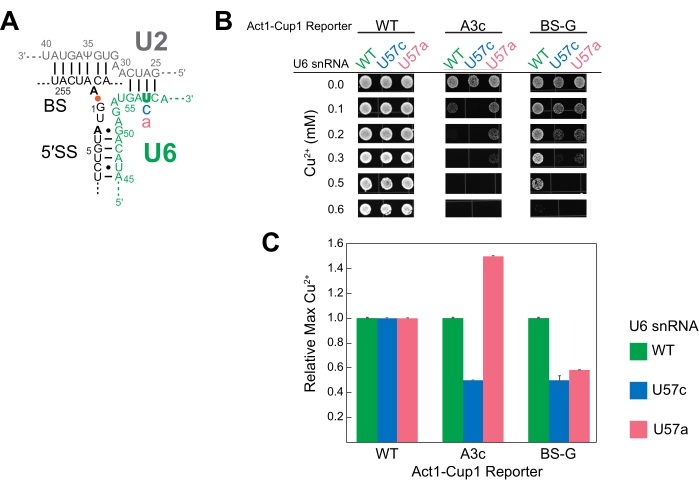
Figure 4: Representative ACT1-CUP1 results monitoring splicing in yeast with splicing component mutations. (A) Schematic of the RNA components of the active site immediately after the first catalytic step. The U2 and U6 snRNAs are duplexed, bringing the 5' SS and BS of the intron in close proximity. The intronic bond between A259 (BS) and G1 (5' SS) is indicated by the orange dot. The locations of the non-consensus sequence substitutions in the ACT1-CUP1 reporter tested in this experiment are indicated in bold black. The mutated position in U6 snRNA (U57) is in bold green, and the substituted bases are in either blue or pink. (B) One possibility for the presentation of ACT1-CUP1 data in a publication includes several images of colonies from relevant Cu2+ concentrations. The WT reporter survived past the copper concentration tested for all three U6 snRNA strains queried. (C) A bar graph comparing the effects per reporter and per U6 snRNA mutant. Normalization is performed for each ACT1-CUP1 reporter by setting the copper tolerance of the U6 snRNA WT to 1 and calculating the ratio for the U57c and U57a mutations. Error bars represent the standard deviation from three replicates. Please click here to view a larger version of this figure.
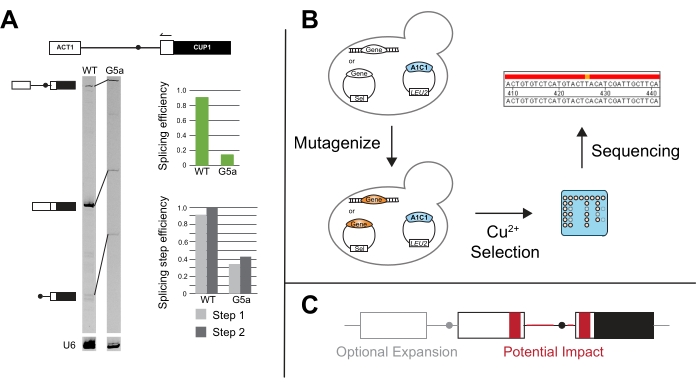
Figure 5: Additional methods to compliment ACT1-CUP1 results. (A) Primer extension is performed via a primer annealing in the 3' ACT1 exon and then elongating into the intron. In this example, the primer is end-labeled with IR700 dye, and the primer extension was performed as described in 20,21. Primer extension of U6 snRNA is performed in the same reaction to serve as a loading control. The 7% 19:1 bis/acrylamide, denaturing gel resolves the pre-mRNA, mRNA, and lariat products after primer extension using a near infra-red gel imaging device as described in van der Feltz et al.22. The intensity of the different bands can be used to measure overall the splicing efficiency as well as distinguish the splicing differences that occur at the first or second step, as quantified with ImageJ44 or other gel band quantifying software. Both the splicing efficiency and the splicing step efficiency are calculated as described in Query and Konarska14. (B) Mutational screens with ACT1-CUP1 reporters can utilize copper selection to identify mutants that impact splicing. The gene(s) of interest can then be sequenced in the resulting strains to determine if the mutation occurred in that gene. (C) Alterations can be made to the reporter outside of the splice sites to monitor splicing changes in relation to intron structure, exonic sequences, number of exons, or nuclear export of unspliced RNA. Regions in grey are optional expansion regions to include on a reporter, and areas in red are regions in which the sequence and structural changes could be tested for their potential impact on splicing using the ACT1-CUP1 assay. Please click here to view a larger version of this figure.
| Reporter name | Intronic region | Sequence | Last viable Cu2+ concentration (mM) | |
| WT | 5’ | GUAUGU | > 2.5 | |
| G1a | 5ʹ | aUAUGU | 0.0133 or 0.0532 | |
| A3c | 5ʹ | GUcUGU | 0.15^,18 or 0.216 | |
| G5a | 5ʹ | GUAUaU | 0.303 or 0.259 | |
| WT | Branch-site | UACUAAC | > 2.5 | |
| C256a | Branch-site | UAaUAAC | 0.1526 or 0.1832 | |
| U257c/a | Branch-site | UACc/aAAC | 0.226 or 0.332 or 0.545 | 0.059,32 |
| A258c/u | Branch-site | UACUc/uAC | 0.826 | 1.045 or 1.646 |
| BS-C | Branch-site | UACUAcC | 0.153 or 0.1832,56 or 0.226 | |
| BS-G | Branch-site | UACUAgC | 0.0516,32 or 0.625 or 0.846 | |
| C260g | Branch-site | UACUAAg | 0.826 | |
| WT | 3’ | UAG | > 2.5 | |
| U301g | 3ʹ | gAG | 0.1518 | |
| A302g/u | 3’ | Ug/uG | 0.0139 | 0.07516 or 0.1824 |
| G303c | 3ʹ | UAc | 0.0532 | |
Table 1: List of common reporters and the Cu2+ concentration lethality reported. There are over 100 citations of Lesser and Guthrie3. While a small selection of these citations was used to create this table, they highlight the general trend of slight to moderate differences between studies in reported copper viability concentrations. In ACT1-CUP1, it is important to compare wild-type protein or RNA to the mutants all with the same yeast strain background using the published concentrations as a guide but anticipating that the observed concentrations may differ. Data gathered from Figure 3 (^) and multiple publications annotated by the following citations3,9,16,18,24,25,26,32,45,46.
Supplemental Table 1: Contents of a single copper plate and an example of calculations done to achieve copper dilutions from 0 mM to 2.5 mM from a 1 M CuSO4 stock. Please click here to download this Table.
Supplemental Table 2: Example of a plating scheme for the 48-pin replicator. Please click here to download this Table.
Supplemental Table 3: Example of viability scored from copper plates incubated for 3 days at 30°C. Please click here to download this Table.
Supplemental Table 4: List of yeast strains used to generate the representative data. Please click here to download this Table.
Supplemental Table 5: List of plasmids used to generate the representative data. U6 snRNA plasmids generated and published in previous research22,47. Please click here to download this Table.
