The experimental workflow for demonstrating intestinal mobility of histidine through absorption of histidine by intestinal villi into the lumen of the inverted sacs is illustrated in Figure 4, Table 1 and Table 2. Three independent experimental setups were performed, and representative data are presented in Figure 6.
Under the given experimental conditions, Histidine estimation using Pauly's reaction followed Lambert Beer's law till 300 µM (Figure 6A). This test for histidine relies on the coupling between amino acids and the diazonium ion in the reagent. Pauly's reagent has sulphanilic acid dissolved in concentrated hydrochloric acid. This acid undergoes diazotization in the presence of sodium nitrate and HCl. The diazonium salt p-phenyldiazosulphonate reacts with histidine molecules under alkaline conditions to form a dark red/orange to a yellow colored complex, which can be spectrophotometrically read at 490 nm (Figure 5)9. It may be noted that this test is given by histidine and tyrosine. The test used in this study is a slightly modified version of the original reaction such that sodium nitrate is used directly at room temperature (~20 °C) rather than using sodium nitrite under chilled conditions9.
The positive Pauly's reaction observed with the sample recovered from inside the inverted sacs illustrates the absorption of histidine across the intestinal villus. Figure 6B shows a correlation between the uptake of histidine and sodium concentrations in jejunal enterocytes. An increase in the absorption of histidine as incubation time increases from 30 to 60 min is indicative of secondary active transport.
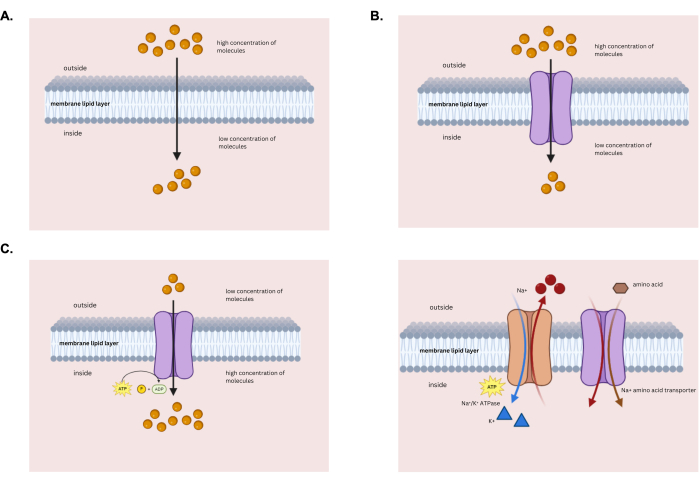
Figure 1: Different types of membrane transport. Schematic showing the various kinds of membrane transport systems. (A) Passive diffusion. (B) Facilitated diffusion. (C) Primary active transport (left) and secondary active transport (right). Please click here to view a larger version of this figure.
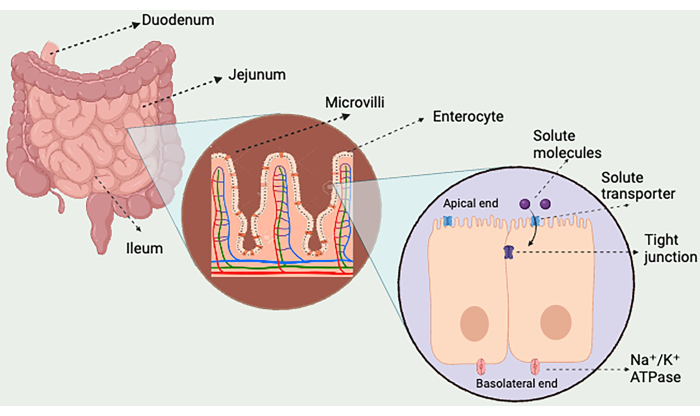
Figure 2: Organization of enterocytes in intestinal villus. This figure focuses on the small intestine and illustrates the three distinct parts, i.e., Duodenum, Jejunum, and Ileum. The small intestine further has some finger-like projections called the villi, which increase the surface area for absorption of ingested food. Microvilli have cells called enterocytes that contain the specific transporters for amino acid absorption. Tight junctions prevent the movement of solutes in between the cells. Please click here to view a larger version of this figure.
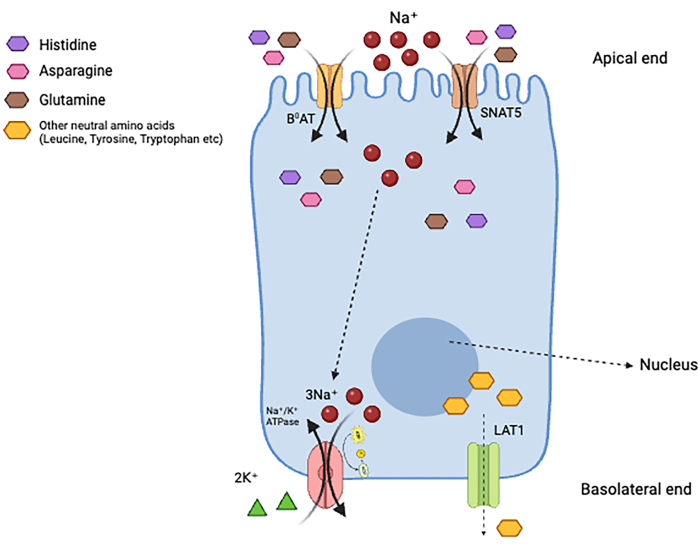
Figure 3: An enterocyte with amino acid transporters. The cartoon represents one enterocyte showing the apical and basolateral ends. The apical end has transporters such as the B0AT and the SNAT5 proteins, and the basolateral membrane contains the LAT1 transporter. Together, these proteins are responsible for the uptake of histidine from the intestinal lumen to the portal circulation. Please click here to view a larger version of this figure.
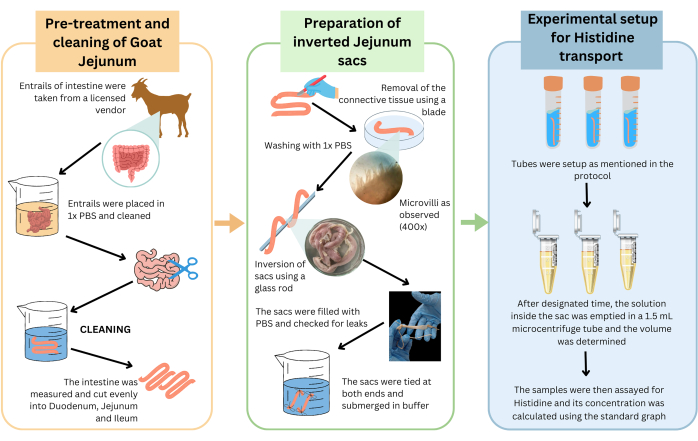
Figure 4: Schematic representation of the protocol demonstrating membrane transport of histidine using goat intestine inverted sacs. Goat intestines were procured, pre-treated, and cleaned as described. Briefly, the jejunum was excised, and inverted sacs were prepared. These sacs were used to set up the experiment to quantify histidine uptake in various salt concentrations. The villi were viewed at 400x magnification using a light microscope. Detailed procedure is provided in the text. Please click here to view a larger version of this figure.
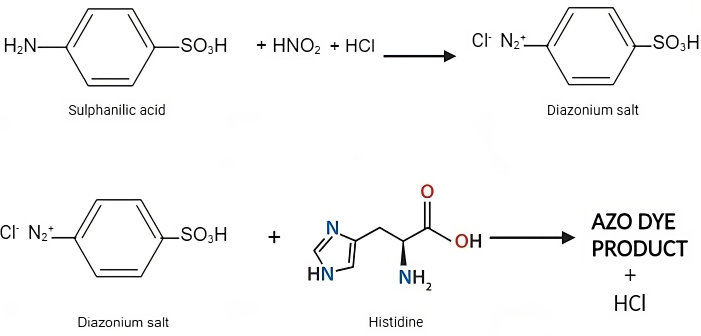
Figure 5: Reaction for Pauly's method. Pauly's test involves the formation of diazonium ions in a reagent containing sulphanilic acid in concentrated hydrochloric acid. Under alkaline conditions, histidine reacts with the diazonium salt, resulting in the formation of a red-colored compound whose absorbance can be measured at 490 nm using a spectrophotometer. Please click here to view a larger version of this figure.
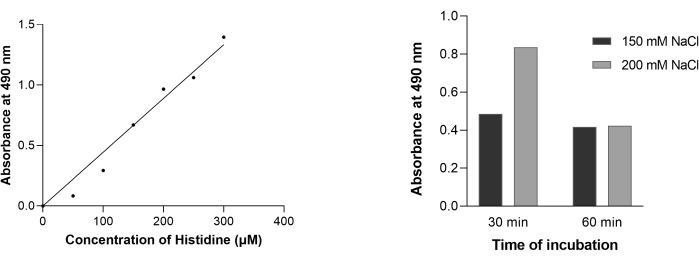
Figure 6: Estimation of histidine by Pauly's method. (A) Standard curve for histidine. Varying concentrations of histidine ranging from 0-300 μM were subjected to Pauly's reaction, and the complex formed was measured at wavelength 490 nm. (B) Transport for histidine in intestinal sacs was followed for 30 min and 60 min in the presence of 150 mM (solid black) and 200 mM NaCl (solid grey) salt solution. The concentration of histidine inside the inverted sacs was estimated using the standard curve (Figure 6A). The values are reported with respect to the control (0 mM NaCl). Three technical replicates were performed, and representative data are presented. Please click here to view a larger version of this figure.
Table 1: Experimental setup for intestinal mobility of Histidine. This table shows the incubation of intestinal sacs in different concentrations of sodium chloride over two time points. Please click here to download this Table.
Table 2: Preparation of standard curve for Histidine. This table shows the different steps and components for Pauly's reaction with known concentrations of histidine ranging from 0-300 μM. Please click here to download this Table.
