To evaluate the pharmacological role of BBR in relieving diabetic tendon injury, cell apoptosis and autophagy activation in foot tendon tissues of DM rats were assessed in the presence or absence of BBR. Figure 1A showed that the protein level of LC3 (an autophagy marker) was decreased in tendons tissues of DM rats compared with control rats, whereas BBR treatment significantly restored autophagy activation. In addition, cell apoptosis was elevated in tendons tissues of DM rats compared with normal tissues, whereas BBR significantly decreased cell apoptosis (Figure 1B). These results demonstrate that BBR increases autophagy activation and decreases cell apoptosis in tendon tissues of DM rats.
Then the role of BBR in regulating tendon fibroblasts viability and apoptosis was assessed in vitro. Figure 2A showed that tendon fibroblast viability was decreased in a dose-dependent manner and that 30 mM of HG was suitable to induce tendon fibroblast damage. The protective role of BBR against tendon fibroblast in the presence of HG was next investigated. As shown in Figure 2B, pre-treatment of tendon fibroblasts with BBR contributed to an increase in cell viability in the presence of HG (30 mM) in a dose-dependent manner. BBR (20 µM) was used to treat tendon fibroblasts in the follow-up study. Furthermore, tendon fibroblast apoptosis was measured in the presence of HG or/and BBR using flow cytometry. Figure 2C,D revealed that the proportion of apoptotic cells was significantly increased after HG treatment, whereas BBR decreased HG-induced tendon fibroblast apoptosis.
Chronic inflammation is a characteristic of type 2 diabetes that is linked to diabetic complications and contributes to insulin resistance. Therefore, the inflammatory factors IL-1β, IL-6, and IL-10 in the HG-treated tendon fibroblasts were measured to determine the effect of BBR on the inflammatory response. The results from qRT-PCR showed that the proinflammatory cytokines IL-1β and IL-6 were significantly increased. In contrast, anti-inflammatory cytokine IL-10 was decreased after HG treatment (Figure 3A–C), indicating that HG triggered a proinflammatory response in tendon fibroblasts. More important, BBR treatment repressed HG-induced proinflammatory response in tendon fibroblasts (Figure 3A–C).
To explore the role of BBR in regulating autophagy activation of tendon fibroblasts, tendon fibroblasts were treated with HG in the presence or absence of BBR, and then autophagy activation was assayed. As shown in Figure 4A, HG treatment decreased autophagy activation in tendon fibroblasts as evidenced by decreased LC3 green puncta, whereas BBR obviously restored the activation of autophagy in HG-treated tendon fibroblasts. Furthermore, the autophagy marker LC3-II and autophagy substrate p62 were assessed using western blot analysis. Figure 4B,C showed that HG decreased LC3  protein expression and enhanced p62 protein expression in tendon fibroblasts, indicating that HG inactivated autophagy of tendon fibroblasts. As expected, BBR restored autophagy activation in HG-treated tendon fibroblasts (Figure 4B,C).
protein expression and enhanced p62 protein expression in tendon fibroblasts, indicating that HG inactivated autophagy of tendon fibroblasts. As expected, BBR restored autophagy activation in HG-treated tendon fibroblasts (Figure 4B,C).
Finally, whether autophagy-mediated the role of BBR in alleviating HG-induced injury of tendon fibroblasts was investigated. To this end, a specific inhibitor of autophagy, 3-MA, was used to inactivate autophagy in the presence of BBR, and then cell viability and proinflammatory cytokines were assessed. As shown in Figure 5A,B, pharmacological inhibition of autophagy increased tendon fibroblast apoptosis compared with BBR treatment alone. Similarly, 3-MA treatment resulted in a significantly decreased tendon fibroblast viability compared with BBR treatment alone (Figure 5C). 3-MA treatment also increased the level of proinflammatory cytokines compared with BBR treatment alone (Figure 5D). Taken together, these data demonstrate that BBR relieves diabetic tendon injury by activating autophagy of tendon fibroblasts.
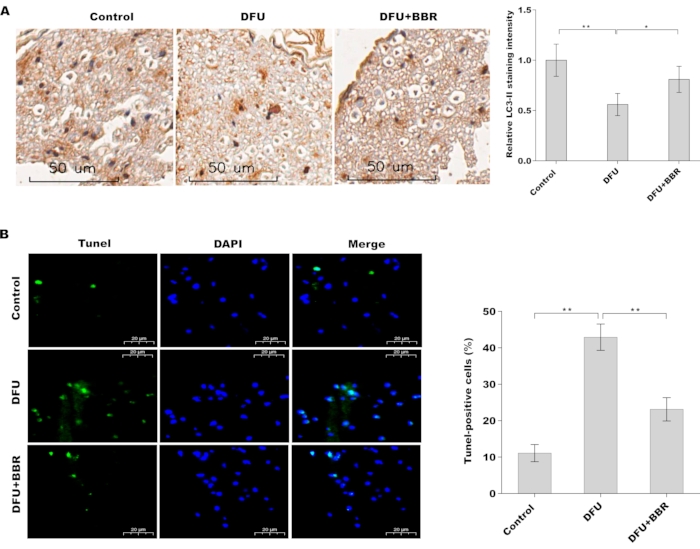
Figure 1: BBR increased autophagy activation and decreased cell apoptosis in tendon tissues of DM rats. (A) Immunohistochemical analysis of LC3 protein expression in foot tendon tissues of control rats (n = 5), DM rats (n = 5), and BBR-treated DM rats (n = 5). Scan bar = 50 µm. Magnification 20x. (B) Cell apoptosis was analyzed using TUNEL staining in foot tendon tissues of control rats (n = 5), DM rats (n = 5), and BBR-treated DM rats (n = 5). Scan bar = 20 µm. Magnification 20x. Please click here to view a larger version of this figure.
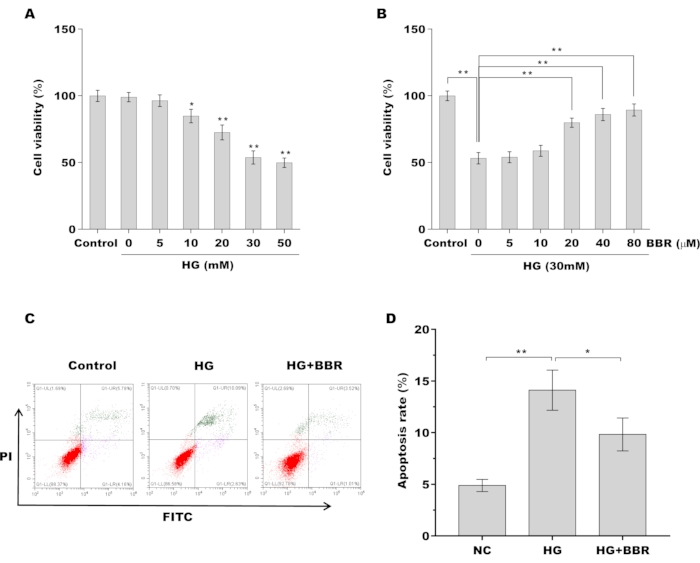
Figure 2: BBR decreased HG-induced apoptosis of tendon fibroblasts (A) Tendon fibroblast viability was assessed using CCK-8 assay after treatment with different doses of HG (0, 5, 10, 20, 30, 50 mM). (B) Tendon fibroblast viability was assessed using CCK-8 assay after treatment with 30 mM of HG in the presence of BBR (0, 5, 10, 20, 40, and 80 µM). (C,D) Cell apoptosis of tendon fibroblasts was assessed using flow cytometry after treatment with HG (30 mM) in the presence or absence of BBR (20 µM). * p < 0.05, ** p < 0.01. Please click here to view a larger version of this figure.
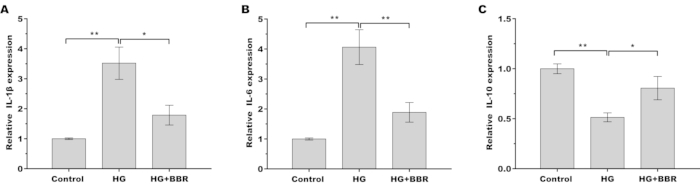
Figure 3: BBR alleviated HG-induced inflammatory response in tendon fibroblasts. (A–C) The mRNA levels of the inflammatory factors (A) IL-1β, (B) IL-6, and (C) IL-10 in tendon fibroblasts were measured by qRT-PCR after treatment with HG (30 mM) in the presence or absence of BBR (20 µM). * p < 0.05, ** p < 0.01. Please click here to view a larger version of this figure.
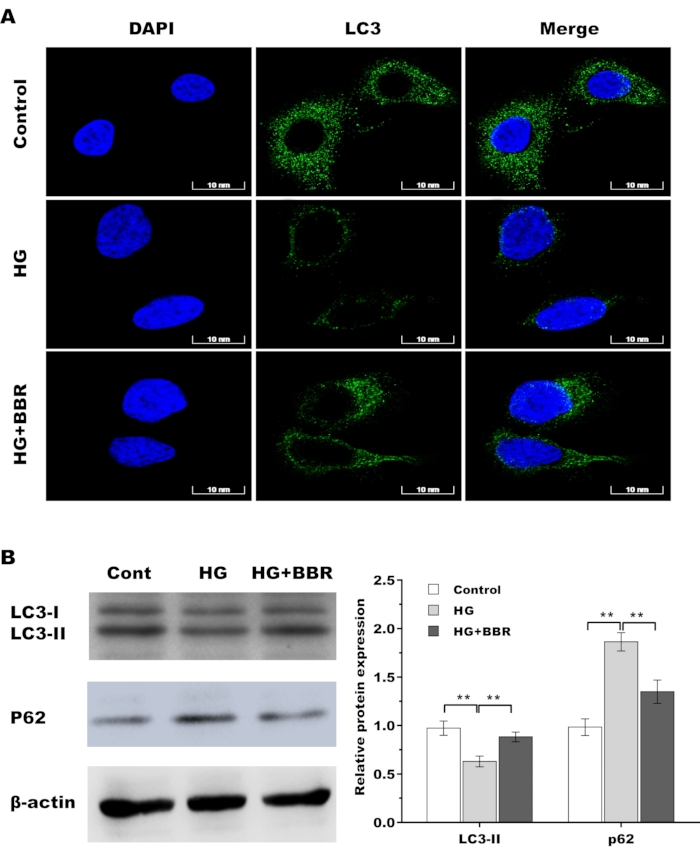
Figure 4: BBR restored autophagy activation in HG-treated tendon fibroblasts. (A) The LC3 level was assessed by immunofluorescence analysis in tendon fibroblasts after treatment with HG (30 mM) in the presence or absence of BBR (20 µM). Scan bar = 10 nm. Magnification 60x. (B) Western blot analysis for the protein levels of LC3 and P62 in tendon fibroblasts after treatment with HG (30 mM) in the presence or absence of BBR (20 µM). * p < 0.05, ** p < 0.01. Please click here to view a larger version of this figure.
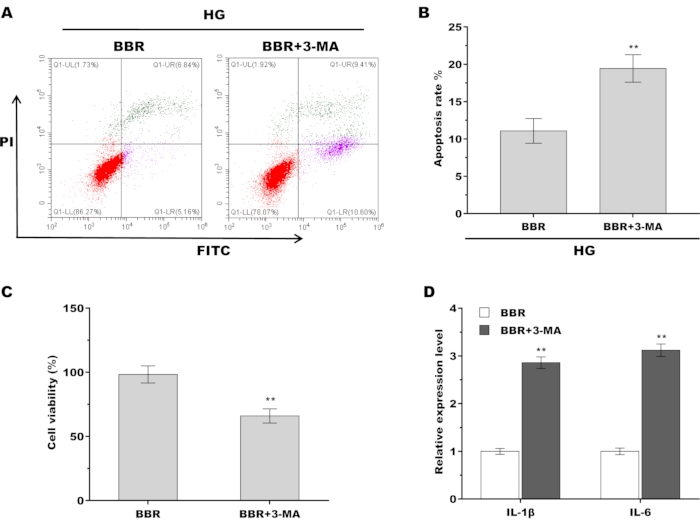
Figure 5: BBR alleviated HG-induced injury of tendon fibroblasts by activating autophagy. (A,B) Cell apoptosis of tendon fibroblasts was assessed using flow cytometry after treatment with BBR (20 µM) alone or BBR plus 3-MA (5 Mm) in the presence of HG (30 mM). (C) Tendon fibroblast viability was assessed using CCK-8 assay after treatment with BBR (20 µM) alone or BBR plus 3-MA (5 Mm) in the presence of HG (30 mM). (D) qPCR analysis of the IL-1β and IL-6 mRNA level in tendon fibroblasts after treatment with BBR (20 µM) alone or BBR plus 3-MA (5 Mm) in the presence of HG (30 mM). ** p < 0.01. Please click here to view a larger version of this figure.
