The CRISPR/Cas9 microinjection protocol described here to generate sand fly mutants was established in a previous publication4. This approach produced highly efficient mutagenesis, as 11 out of 540 individuals survived the procedure, of which 9 were mutant. When designing guides for CRISPR/Cas9 mutation, a critical first step is to sequence the region around the area to be targeted. The template for sequencing should be from the strain that is going to be used as a source of embryos for injection. It is risky to rely only on published genome sequences to design guides. It is not unusual to have differences between published genome and pre-guide design sequences. In some cases, guides designed from published genome sequence are not present when the region around the target gene is sequenced (Harrell, personal observation). In addition, the sequence may include Single Nucleotide Polymorphisms (SNPs) that could be mistaken as real CRISPR edits during genotyping. Therefore, confirming the sequence for the target region is critical for the rest of the protocol to succeed.
Successful genetic modification of insects by microinjection depends mainly on two critical aspects: delivery of materials (protein, plasmid, or mRNA) at the appropriate time in development with as little damage to the embryos as possible; and rearing of robust, healthy insects that will survive the procedure and produce offspring. The initial phase of this procedure, described in Figure 2, starts with blood-feeding of the females from the wt colony and proceeds to embryo collection and microinjection five days later. The second phase consists of rearing the injected embryos until adulthood, making the appropriate crosses and identifying and isolating mutations of interest.
Embryos for injection are collected for 30 – 60 min so that the relative age in hours post oviposition can be determined. Embryos are then allowed to develop for 3 h before the start of the injections. This time window of insect development allows the embryos to survive the injections. After this aging period, the embryos are collected onto a prepared injection slide and the embryos are rolled into place against a coverslip edge using a fine brush. The final configuration is shown in Figure 3A. It is important to wet the filter paper enough so that a meniscus forms at the coverslip edge where the embryos sit. Too much water and the embryos will be pushed away from the coverslip edge. Too little will cause the embryos to be drawn under the coverslip. The embryos need to be kept damp, otherwise the embryo membranes become difficult to inject. The coverslip edge acts as a back stop, allowing the needle to penetrate the embryos when pressed against the edge. It is the combination of a sharp needle and a backstop that allows for the needle to penetrate smoothly.
As with most successful microinjection protocols, a good microinjection needle suited to the embryos being injected is important. Good, sharp needles are defined as needles that easily penetrate the embryo without allowing the material to escape post-injection. Good penetration is evident when the needle slips into the embryo, causing little to no indentation of the embryo membrane during penetration and no material leaks from the embryo after the needle is withdrawn. Good sharp needles are produced using needle puller settings that produce a needle that comes to a fine point (Figure 1A). The pulled needle should not have a taper that is too long. Otherwise, the lumen of the needle becomes very narrow for a major portion of the taper (Figure 1B), and it becomes difficult to get the injection pressure high enough to force the material through the needle. In this protocol, beveled needles were used. The process for pulling and beveling needles is described in Meuti and Harrell19. In the case of sand fly embryos that develop over a long period of time, it is particularly critical to have sharp needles that minimize damage to the embryo, thus preventing material from leaking at the injection site. When embryoplasm leaks from an embryo post-injection, this material is a rich medium for mold and fungal growth. In embryos that develop over a shorter period of time the embryo can develop and hatch before the mold becomes an issue. Any embryos that are visibly leaking embryoplasm after injection should be removed.
During injections, it is important to add water as needed by wetting a fine paintbrush and touching it to the filter paper, repeating as necessary until the meniscus of water is just at the base of the embryos. Attention to relative humidity on the injection day needs to be noted. On low humidity days, more water will need to be added. Once injections are completed, a little extra water is added to the filter paper so that the coverslip slightly floats, making it easier to remove. Once the coverslip has been removed, the filter paper with injected embryos can be blotted, so that the filter paper is just barely damp. Embryos can then be transferred to a moist plaster pot for hatching using a fine paintbrush. At this stage, embryos are very fragile so the process has to be undertaken very carefully. It is also important to prevent the embryos from touching each other to be able to remove moldy embryos and limit the spread of fungal contamination.
Post-injection, the G0 injected embryos are kept on plaster pots as per normal rearing procedures. Until they hatch, injected embryo pots should be checked once a day to remove unhealthy embryos. Figure 4A presents the expected timeline of G0 individual rearing from the day of egg-laying and injection to adulthood. Figure 4B shows examples of G0 embryos that are healthy and should be retained; or damaged, contaminated by fungus, desiccated, or deformed and should be discarded. The G0 injected individuals are supposedly mosaic for mutant alleles. A method for identifying potential mutant alleles should be designed, such as a PCR-assay of the region surrounding the expected cutting site(s). Figure 4C presents an example of a PCR-assay showing a mosaic G0 individual, exhibiting a PCR product additional to the expected wt product, representing a mutant allele.
Once the adults emerge, the G0 females developing from G0 injected embryos are crossed with wt males, allowed to lay eggs, and are genotyped later. Only the tubes containing a G0 fly showing evidence of mutation from the genotyping method of choice are retained. The flies from the next generations (G1 females) are crossed either with wt males or individually between siblings (from G2). These crosses are allowed to lay eggs and then genotyped by the method of choice. The last step is repeated until homozygous mutant males and females are obtained, establishing a homozygous mutant line. A schematic representation of the appropriate succession of crosses to establish a mutant line is given in Figure 5A. Figure 5B shows an example of a genotyping PCR allowing the identification of a homozygous mutant sibling cross.

Figure 1: Microinjection needles. A. Good needle. B. Needle with extreme taper not good for injections. Please click here to view a larger version of this figure.
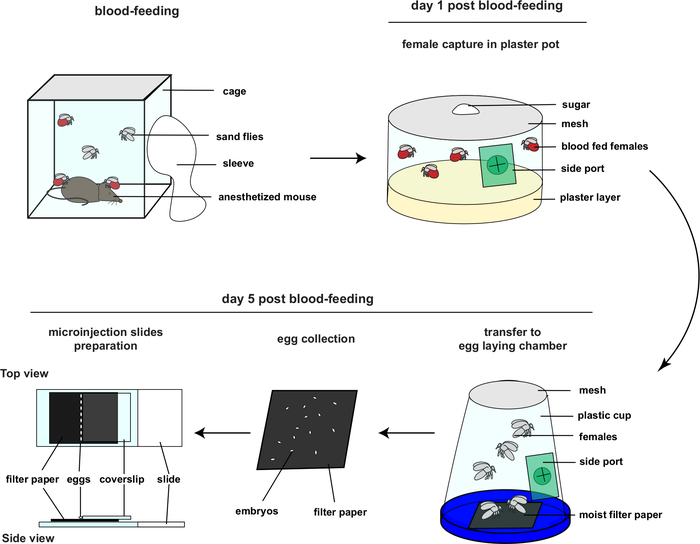
Figure 2: Overview of embryo collection and micromanipulation (This figure had been adapted from4). Please click here to view a larger version of this figure.
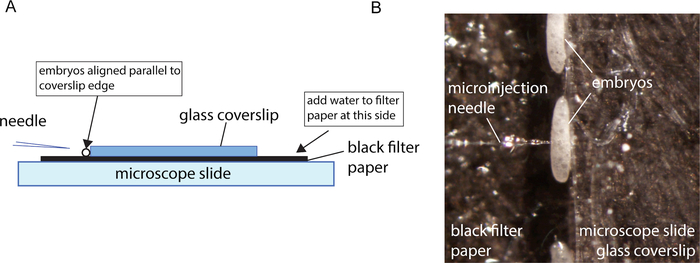
Figure 3: Microinjection setup. A. Schematic representation of the microinjection set-up. B. Close-up of aligned embryo for microinjection. Please click here to view a larger version of this figure.
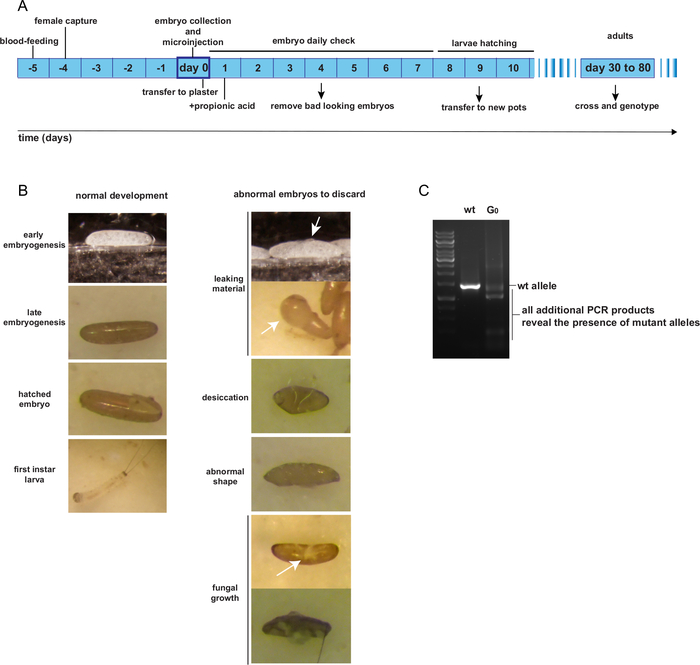
Figure 4: Rearing and identification of mosaic G0 adult individuals. A. Expected time schedule from embryo microinjection to G0 adulthood. B. Examples of good and bad looking embryos post-injection. C. PCR genotyping of one transformed G0 mosaic individual. Please click here to view a larger version of this figure.
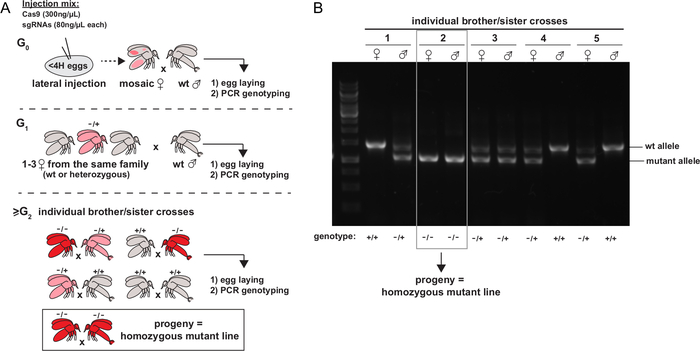
Figure 5: Identification and isolation of mutations of interest. A. Schematic representation of the experimental strategy to isolate mutant alleles and establish homozygous mutant stocks (figure from 4). The color represents the presence of cells carrying mutant alleles, at the heterozygous (pink) or homozygous (red) state. B. Example of a PCR screening strategy for genotyping individual sibbling crosses of sand flies. Please click here to view a larger version of this figure.
