Figure 2 shows representative live-imaging results of this protocol. When the protocol is successful, Dpp-GFP can be seen as a stripe down the center of the wing disc with nuclei visible as non-fluorescent circles within the Dpp-GFP region (Figure 2). Dpp-GFP release is visible as fluorescent puncta that appear and disappear. We have observed Dpp-GFP fluorescence appearing and disappearing in the cell bodies and far from the cell bodies. Dpp signaling is dependent on actin-based filopodia-like structures called cytonemes that extend far from the cell body22,23. Therefore, it is likely that Dpp-GFP puncta that are visualized far from the Dpp-producing cell bodies in these presented videos are likely in cytonemes or at cytoneme “synapses”15. These puncta are most apparent close to the D/V boundary, which can be seen as the gap in the Dpp-GFP stripe.
Figure 3 shows a suboptimal result. In the method described here, the wing discs are mounted so that the peripodial side faces away from the microscope slide so that the disc is imaged from the peripodial side (as shown in Figure 1B). If the wing disc is imaged from the reverse side, that is, peripodial side toward the microscope slide, the Dpp-GFP fluorescence will appear out of focus and resolution will be poor, resulting in the suboptimal result shown in Figure 3. Step 4.6 is therefore critical to ensure that the wing disc is lying flat in the correct orientation before imaging to avoid this result.
Figure 4 shows a time series of images of a Dpp>GFP (dpp-GAL4; UAS-GFP) third instar larval wing disc. When GFP is not fused to Dpp, we do not observe the appearance of puncta outside of the cell bodies. This control is important for the conclusion that Dpp-GFP behaves differently than GFP alone.
Figure 5 shows a time series of images of a dpp-GAL4 driver alone third instar larval wing disc. Images were taken with the same microscope acquisition settings as all of the other samples. These images show that fluorescent puncta do not appear when Dpp-GFP is not expressed.
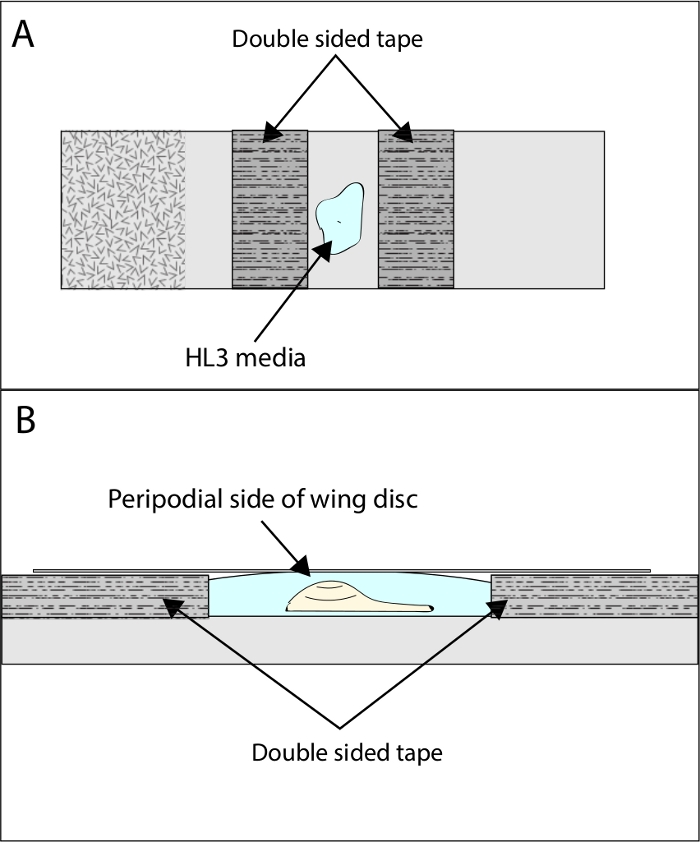
Figure 1: Illustration of wing disc mounting setup. To image Dpp-GFP release, the dissected wing discs should be mounted as shown. (A) Two pieces of double-sided tape are placed widthwise across the microscope slide with the wing disc mounted in modified HL3 media between the tape. (B) Side view of a correctly mounted wing disc. The disc is oriented so that the peripodial side of the wing pouch faces upward towards the coverslip. Please click here to view a larger version of this figure.
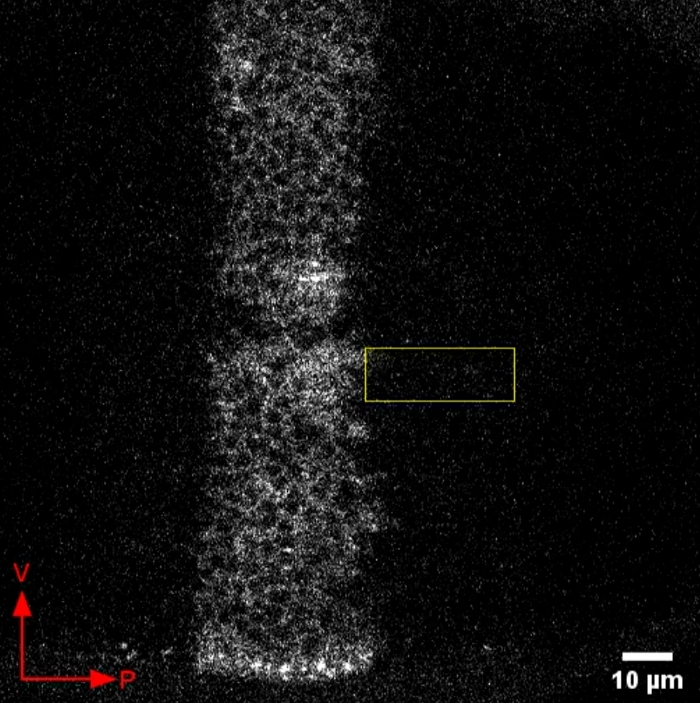
Figure 2: Representative video of Dpp-GFP release in the wing disc. Dpp-GFP was expressed in the wing disc using the UAS-GAL4 system. Shown here is a representative video of live imaging results of Dpp-GFP release. Cell nuclei can be seen as circular gaps in fluorescence. Secreted Dpp-GFP can be seen as bright puncta. Due to the contrast, these bright puncta are easiest to observe outside of the region that includes the Dpp-producing cell bodies. Such an area is indicated by a rectangle that disappears after the first few frames. Ventral and posterior directions of the wing disc are indicated by the arrows. Please click here to download this video.

Figure 3: Suboptimal result of Dpp-GFP release imaging. Example of a suboptimal result which occurs if the wing disc is orientated peripodial side face-down. Due to the incorrect orientation of the wing disc pouch Dpp-GFP is not in focus, clear nuclei cannot be seen, and resolution remains poor no matter how the parameters of the microscope are adjusted. Ventral and posterior directions of the wing disc are indicated by the arrows. Please click here to download this video.
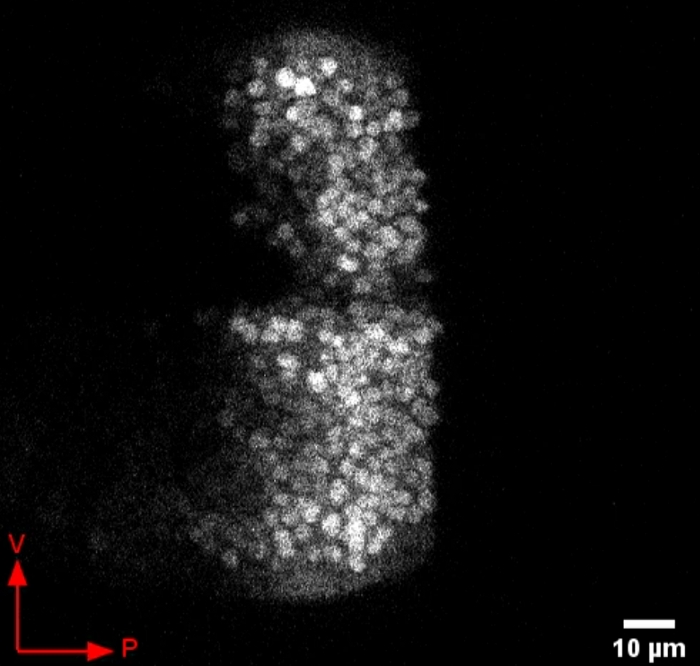
Figure 4: Representative video of GFP expression in Dpp-producing cells of the wing disc. GFP expression appears brightly in the cell bodies and does not appear as bright puncta released in the periphery. Ventral and posterior directions of the wing disc are indicated by the arrows. Please click here to download this video.

Figure 5: Representative video of dpp-Gal4 third instar wing discs without the expression of Dpp-GFP or GFP. If Gal4 cannot drive expression of GFP or Dpp-GFP due to lack of UAS-GFP or UAS-Dpp-GFP, no fluorescence is observed. Please click here to download this video.
