Enzymatic dissociation of DSM pieces provides healthy freshly isolated DSM cells routinely used in functional and molecular studies such as: patch-clamp electrophysiology and immunocytochemistry. Figure 1 summarizes the dissection steps and visualizes the setups employed for temperature control of enzymatic treatment steps. Figure 2 illustrates bright-field images of DSM cells obtained from three human urinary bladder specimens each from a different patient-donor. Healthy single DSM cells are characterized by spindle-shaped morphology, crisp well-defined edges, a well-defined halo around the cell, and semi-contractile (serpentine-like) appearance when viewed under the microscope (see DSM cells demarked by white arrows in Figure 2). They also respond to contraction stimulating agents such as the muscarinic agonist carbachol or high K+ (60 mM) applications. DSM cells show positive immunoreactivity for α-smooth muscle actin confirming their identity (Figure 3).
DSM cells are ideally suited for patch-clamp electrophysiological investigations of ion channel properties. Here, we describe the amphotericin-B perforated patch-clamp recording method using pipette and extracellular solutions (Table 2) to optimally record voltage-step induced cation channels. Under the specific conditions employed, blockade of Kv and L-type Ca2+ currents with Cs+/TEA+ and nifedipine, respectively, ensured the elimination of contribution of these ionic components to the whole-cell voltage-evoked currents. Figure 4 and Figure 5 show, respectively, the experimental steps of the amphotericin-B perforated patch-clamp method and representative whole-cell currents measured either with a voltage-step induced or a ramp protocol in three different human DSM cells, each from a different patient donor. Note that the recordings display a certain degree of variability in terms of current amplitudes and outward rectification. Additional experiments revealed that 9-phenanthrol, a TRPM4 channel inhibitor, effectively and reversibly inhibited human DSM cation currents at negative and positive voltages (Figure 6). The 9-phenanthrol-sensitive current component illustrates a stronger inhibition at positive voltages and outward rectification (Figure 6C).
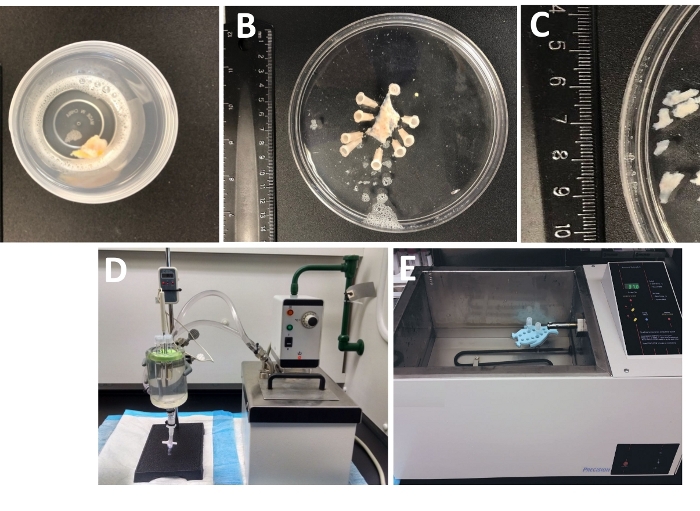
Figure 1: Summary of dissection steps resulting in the preparation of detrusor smooth muscle (DSM) pieces and setup used for enzymatic dissociation. Shown are images of: (A) a whole thickness human urinary bladder specimen provided from an open bladder surgery as extraneous surgical material in ice-cold DS, (B) the same preparation after pinning with a partially dissected DSM layer, (C) DSM pieces of variable dimensions cut out from the DSM layer ready for enzymatic digestion (smaller pieces) or other experimental investigations (larger pieces), (D, E) alternative setups used for enzymatic digestion of DSM pieces consisting of either (1) a temperature-controlled circulating water bath connected via tubing to a large glass tissue chamber filled with water, a rubber holder for tubes, plastic tubes containing DSM pieces and enzyme solutions prepared in dissection/digestion solution (DS, either DS-P or DS-C, Table 2) and a temperature probe linked to a display allowing for continuous monitoring (D), or (2) a large water-filled temperature-controlled bath containing a holder and tubes with DSM pieces and enzyme solutions (E). Please click here to view a larger version of this figure.
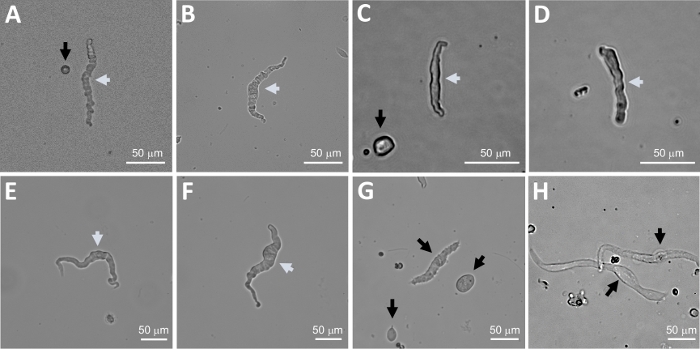
Figure 2: Representative bright-field images of human freshly isolated DSM cells obtained using the sequential papain-collagenase digestion method. (A-F) Displayed are images of viable, physiologically active DSM cells considered suitable candidates for attempting perforated patch-clamp recordings. (G, H) Images of non-viable or over-digested cells; such cells were avoided for patch-clamp experiments. White and black arrows in panels (A-H) point to DSM cells considered viable and non-viable, respectively, for attempting patch-clamp recordings. Note that the black arrows in panels (A, C, and G) point to cell fragments (circular pieces) or small cells lacking DSM morphology and in (H) the cells appear pale and dilated. Images are from three different urinary bladder specimens (A and B : patient-donor source one, C and D : patient-donor source two, and E-H : patient-donor source three). Please click here to view a larger version of this figure.
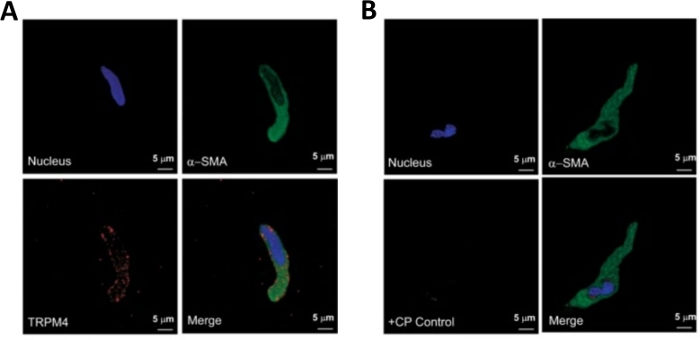
Figure 3: Expressions of transient receptor potential melastatin type 4 (TRPM4) channel and α-smooth muscle specific actin immunoreactivities in single human DSM cells by immunocytochemistry analysis. (A) Shown are confocal images showing immunocytochemical detection of TRPM4 channel protein expression in a human DSM cell. Red staining (bottom left) indicates TRPM4 channel proteins; blue (DAPI) staining detects cell nuclei (top left); green staining indicates α-smooth muscle actin (α-SMA, top right); the merged image (bottom right) illustrates the overlap of all three images. (B) Confocal images illustrating attenuation of immunocytochemical detection of TRPM4 channel protein expression in the presence of a TRPM4-specific competing peptide (CP) in isolated human DSM cells. Blue (DAPI) staining indicates cell nuclei (top left); green staining is for α-smooth muscle actin (α-SMA, top right); the merged image (bottom right) illustrates the overlap of all three images. The results were verified in four separate experiments using DSM whole tissue or multiple DSM cells isolated from four patients. Images are from Hristov et al. (2016)22 and used with permission. Please click here to view a larger version of this figure.
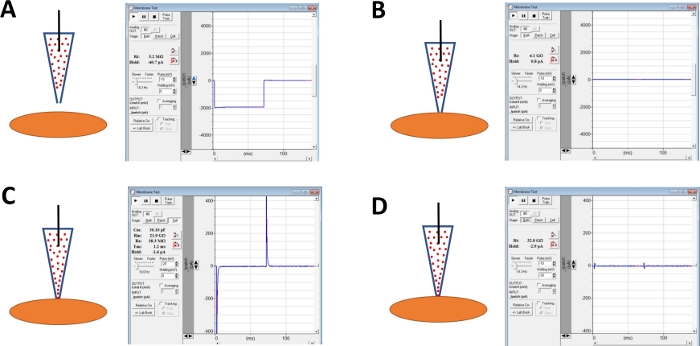
Figure 4: Schematic illustration of steps involved in giga-seal formation and amphotericin-B perforation of human DSM cells. Illustrated are spatial positions of an amphotericin-B containing pipette and a DSM cell along with associated responses for membrane tests obtained in the commercial acquisition software (Table of Materials) by altering voltage steps (either -10 or -20 mV in this example) determining resistance. Configurations are: (A) prior to the cell approach with an electrode, (B) after giga-seal formation obtained by positioning the amphotericin-B containing pipette (amphotericin-B represented by red dots) onto the cell surface and applying negative pressure, (C) the on-cell configuration shown ~45 min after giga-seal formation, at this time-point amphotericin-B has diffused down the pipette and its molecules have inserted into the plasma membrane at the tip of the electrode forming cation permeable pores, and (D) the same configuration as in (C) but with capacitance transients canceled out using dials for whole-cell capacitance and series resistance on the amplifier. Please click here to view a larger version of this figure.
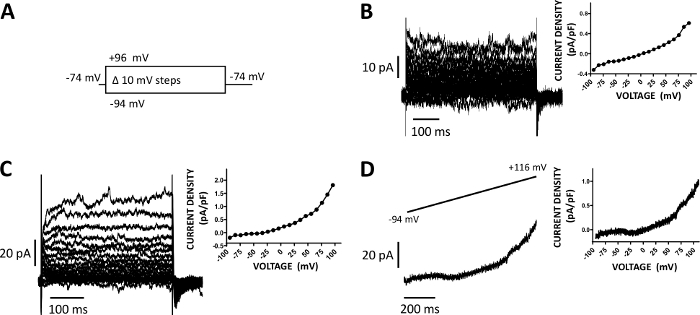
Figure 5: Whole-cell cation currents recorded with amphotericin-B perforated patch-clamp method in human DSM cells. (A) A diagram of the voltage-step protocol illustrates a holding potential of -74 mV and voltage-steps of 400 ms duration from -94 to +96 mV carried out in 10 mV increments and then returned to -74 mV. (B, C) Representative current traces along with current density-voltage plots from two different human DSM cells, each from a different urinary bladder specimen/patient-donor obtained with the voltage-step protocol described in (A). (D) An example of a current trace obtained with a ramp protocol (graphically represented in the top inset as voltage change from -94 to +116 mV over a 1 s duration at 0.21 mV/ms, holding potential was -94 mV). On the right in panels (B-D), the graphs display the current density-voltage relationship for each DSM cell recorded. Please click here to view a larger version of this figure.
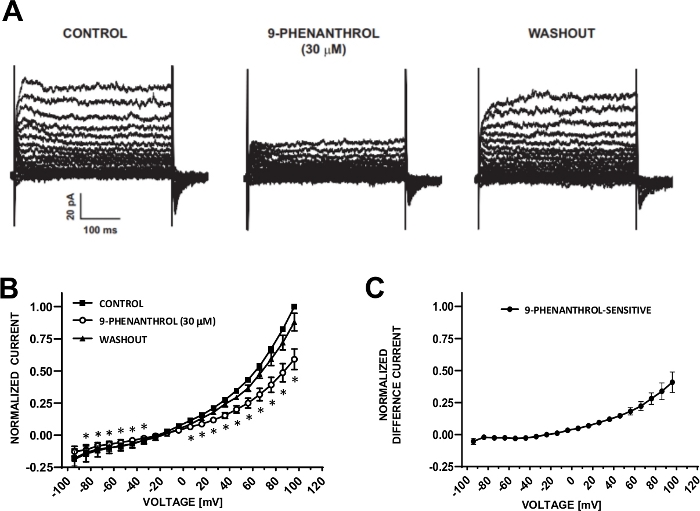
Figure 6: TRPM4 channel blocker 9-phenanthrol-mediated inhibition of voltage-step induced cation currents in human DSM cells. (A) Shown are representative currents measured with the voltage-step protocol described in Fig. 5A for control, 9-phenanthrol, and washout. (B) Summary of normalized responses versus voltage for control, 9-phenanthrol, and washout in seven DSM cells (from seven different patient-donors). (C) Difference current for 9-phenanthrol-sensitive component obtained by subtracting the values in the presence of 9-phenanthrol (30 µM) from those of the control shown in (B). Data in (B) and (C) are displayed as means with error bars for SEM, * depicts significance (p<0.05, paired Student's test) for the comparison of control vs 9-phenanthrol at each voltage. Panels (A) and (B) have been reproduced from Hristov et al. (2016)22 and used with permission. Please click here to view a larger version of this figure.
| Species | Procedure Details | 参考文献 | |
| Guinea pig | DSM pieces rinsed with Ca2+-free medium (in mM: 100 NaCl, 10 KCl, 1.2 KH2PO4, 5 MgCl2, 20 glucose, 50 taurine, measured pCa=6 (or 1 mM) and then cut into pieces and treated typically 90-120 min (4 periods of 30 min) with enzyme medium (in mM: 130 KOH, 20 taurine, 5 pyruvate, 5 creatine, 10 mM HEPES, adjusted with methansulfonic acid to pH 7.4, 1 mg/ml collagenase, 0.2 mg/ml pronase E, 1 mg/ml fatty acid free albumin, pCa=4.2 (63 mM) or 3.7 (200 mM). Single DSM cells were stored in Kraft-Bruhe (KB)-medium (in mM: 85 KCl, 30 K2PO4, 5 MgSO4, 5 Na2ATP, 5 K-pyruvate, 5 creatine, 20 taurine, 5 beta-OH-butyrate, 1 mg/ml fatty acid free albumin, adjusted with KOH to pH 7.2). Alternative method: DSM pieces rinsed for 10 min in a Ca2+-free medium (in mM): 140 NaCl, 5 KCl, 1.2 MgCl2, 10 glucose, 20 taurine, 5 HEPES, adjusted with NaOH to pH 7.4). Then DSM pieces, incubated in the same Ca2+-free medium complemented with 5 mg% collagenase, 2 mg% pronase and 100 μM CaCl2 for 2×20 min stirring. |
Klockner & Isenberg (1985)13,34 Klockner & Isenberg (1986)35 Schneider et al. (1991)10 Bonev & Isenberg (1992)9 Weidelt & Isenberg (2000)36 Moore et al. (2004)14 | |
| Guinea pig, Landrace pig, and human | DSM pieces pre-incubated for 5 min in Ca2+-free Krebs solution then cut into pieces and enzymatically digested in Ca2+-free Krebs solution containing 0.5-2 mg/ml collagenase type I and 0.1-0.5 mg/ml pronase at 36°C for 20-30 min constantly stirred. In some cases, digested pieces were further agitated by a blunt tipped pipette or by spinning until yielding cells. Isolated cells were stored in modified Krebs solution (described in Klockner & Isenberg13) and were normally used within 3 h. The composition of the Krebs solution was (mM): 140 Na+, 6 K+, 2 Ca2+, 1.2 Mg2+, 152.4 Cl–, 10 glucose, 10 HEPES, pH 7.35-7.4 with Tris. For Ca2+-free solution, Ca2+ and Mg2+ were omitted from the Krebs solution. | Inoue & Brading (1990)37 Inoue & Brading (1991)38 Nakayama & Brading (1995)39,40 | |
| Human | DSM pieces placed in Ca2+-free HEPES Tyrode's solution (in mM: 105.4 NaCl, 20.0 or 22.3 NaHCO3, 3.6 KCl, 0.9 MgCl2, 0.4 NaH2PO4, 19.5 or 4.9 HEPES, 5.4 or 5.5 glucose, 4.5 or 5.5 Na-pyruvate) and cut into DSM pieces. DSM pieces soaked in an enzyme solution (Ca2+ free HEPES solution with 0.7 mg/ml collagenase type I, 0.7 mg/ml papain, 1 mg/ml albumin) overnight at 4°C. The strips were then heated at 36.5°C for 15 to 30 min, washed and gently triturated in fresh solution. Isolated cells that were stored in Ca2+ containing HEPES Tyrode's solution or used immediately for experiments. | Montgomery & Fry (1992)24 Gallegos and Fry (1994)41 Fry et al. (1994)42 Sui et al. (2001)43 Wu et al. (2002)44 | |
| Guinea pig | DSM cut into pieces in PSS (in mM: 137 NaCl, 5.4 KCl, 2 MgCl2, 2 CaCl2, 0.42 KH2P04, 4.17 NaHCO3, 10 glucose, 10 HEPES, pH 7.4 with NaOH). DSM pieces placed for 10 min in in the following digestion solution (in mM: 80 Na-glutamate, 55 NaCl, 6 KCl, 10 HEPES, 11 glucose, 2 MgCl2, and 0.2 CaCl2) and then transferred to a vial containing the same solution but with 1 mg/ml collagenase 2, 1 mg/ml trypsin inhibitor (sometimes omitted), 1 mg/ml fat-free bovine albumin, for ~ 70 min at 35°C or ~60 min at 37°C. Single DSM cells were obtained by trituration through a Pasteur pipette in the same solution without calcium and enzymes. After trituration, Ca2+ (1 mM) was added and the cells were stored at 4°C. The cells were always used on the same day. | Bonev & Nelson (1993)53,54 Heppner et al. (1997)26 Petkov et al. (2001)47 Shieh et al. (2001, 2007)48,49 | |
| Guinea pig, mouse, rat, and human | Protocol utilizes a two-step enzymatic dissociation treatment following sharp dissection in Ca2+-free digestion solution (in mM: 80 Na-glutamate, 55 NaCl, 6 KCl, 10 HEPES, 11 glucose, and 2 MgCl2). First, DSM pieces were treated for 25-45 min at 37°C with 1-2 mg/ml papain, 1 mg/ml dithioerythreitol and 1 mg/ml bovine serum albumin in dissociation solution (in mM: 80 monosodium glutamate, 55 NaCl, 6 KCl, 2 MgCl2, 10 HEPES, and 10 glucose, adjusted to pH 7.3 with NaOH) and then DSM pieces transferred to digestion solution containing 1-5 mg/ml collagenase XI (Sigma) or collagenase type 2, 1 mg/ml bovine serum albumin, 0 or 1 mg/ml trypsin inhibitor and 100 μM Ca2+, for 6-30 min. Following incubation, the digested tissue was washed several times in digestion solution without enzymes and Ca2+ and then gently triturated to yield single smooth muscle cells. | Petkov et al. (2001)50 Thorneloe & Nelson (2003)51 Thorneloe & Nelson (2004)33 Petkov & Nelson (2005)27 Hristov et al. (2008)52 Layne et al. (2010)53 Hristov et al. (2011)15 Xin et al. (2012)54 Parajuli et al. (2012)25 Malysz et al. (2013)29 Parajuli et al. (2013)31 Lee et al. (2013)55 Malysz et al. (2014)23 Smith et al. (2013)19,20 Hristov et al. (2016)22 Lee et al. (2017)56 Yarotskyy et al. (2018)57 | |
Table 1: Summary of enzymatic approaches used for isolating single DSM cells from urinary bladders of various species.
| Solution Type | Composition (in mM) |
| DS (Dissection/Digestion Solution) | 80 Na-glutamate, 55 NaCl, 6 KCl, 10 HEPES, 2 MgCl2, and 11 glucose, pH adjusted to 7.4 (with 10 M NaOH) |
| DS-P (Papain-containing DS) | DS containing 1-2 mg/ml papain, 1 mg/ml dithiothreitol and 1 mg/ml bovine serum albumin |
| DS-C (Collagenase-containing DS) | DS solution containing 1-2 mg/ml collagenase type II, 1 mg/ml bovine serum albumin, 0 or 1 mg/ml trypsin inhibitor and 100-200 μM Ca2+ |
| P (Pipette) | 110 CsOH, 110 aspartic acid, 10 NaCl, 1 MgCl2, 10 HEPES, 0.05 EGTA, and 30 CsCl,pH adjusted to 7.2 with CsOH, and supplemented with amphotericin-B (300-500 μg/ml) |
| E (Extracellular) | 10 tetraethylammonium chloride (TEA), 6 CsCl, 124 NaCl, 1 MgCl2, 2 CaCl2, 10 HEPES, and 10 glucose, pH adjusted to 7.3-7.4 with NaOH or CsOH, and 0.002-3 (2-3 mM) nifedipine |
Table 2: Compositions of dissection/digestion solution (DS), and pipette and extracellular solutions used in perforated patch-clamp experiments.
