This procedure has been successfully implemented to isolate tracheal brush cells for RNA sequencing3. After isolation of the trachea and digestion of the tissue with a 2-step protocol (Figure 1), cells were collected and stained with fluorescently-labeled CD45 and EpCAM after exclusion of dead cells with PI. After gating out doublets based on forward and side scatter characteristics, we defined brush cells as low/negative for CD45, positive for EpCAM and positive for eGFP (Figure 2A). Brush cells represented ~0.16-0.42% of CD45low/neg cells by flow cytometry and accounted for 250-600 cells per trachea (Figure 2B). We compared these counts to an estimate of the number of ChAT-eGFP positive cells in whole tracheal mounts from ChAT(BAC)-eGFP mice based on our published extensive immunohistological evaluation of tracheal brush cells3 and illustrated here (Figure 3). Our previous studies of tracheal brush cells in wild type, ChAT(BAC)-eGFP mice and Il25F25/F25 mice suggested that cholinergic brush cells are 90% overlapping with DCLK1+ and IL25+ cells and account for 600-1000 brush cells per trachea3. Notably, this number is lower than the number of cholinergic brush cells reported by other groups10. As chemosensory cell numbers are altered by exposure to microbial metabolites and protozoa5,14, these numbers might reflect a variability in interinstitutional microbiota. Therefore, we would suggest estimating the number of brush cells by fluorescence microscopy to gauge the expected number of isolated brush cells by flow cytometric sorting.
While establishing our protocol, we attempted several previously published methods of isolation of chemosensory cells (Figure 4). When we incubated minced trachea with 650 U/mL collagenase IV and 1.84 U/mL dispase supplemented with DNase I for 45 min, a combination frequently used to obtain single cell suspensions from lung homogenates15, we achieved a single cell suspension of epithelial cells, but we recovered only few brush cells (Figure 4A). Tuft cells in the intestine have been successfully isolated using 2.5 mM EDTA and 0.75 mM Dithiothreitol (DTT) with DNase I for 20 min to dislodge the epithelial cells followed by digestion of the released epithelial cells with 0.01 U/mL dispase with DNaseI for 10 min6. In our hands, using this procedure also led to suboptimal tracheal brush cell recovery (Figure 4B). A protocol for successful isolation of tracheal brush cells for RNA analysis was described by Krasteva et al.10. Following this protocol, we incubated minced trachea with 35 U/mL papain in Tyrode buffer for 45 min and did not achieve good brush cell recovery (Figure 4C). Rock et al. described a method of digestion of tracheal epithelium for basal cell isolation with two steps: separation of the epithelium from the mesenchymal layer with high dose dispase (16 U/mL) at room temperature followed by incubation of the stripped epithelium with 0.1 % trypsin and 1.6 mM EDTA for 30 min at 37 °C16. Following these steps led to a better recovery of brush cells, but the numbers achieved per mouse were insufficient for in depth functional analyses (Figure 4D). To optimize the protocol, we decided to combine the last two approaches. By separating the tracheal epithelium with high dose dispase, we aimed to allow for better access of papain to tracheal brush cells. Thus, using 16 U/mL of dispase at room temperature on whole trachea to separate the epithelium and subsequent incubation of the epithelium with 26 U/mL papain in Tyrode buffer led to the best brush cell recovery (Figure 4E).
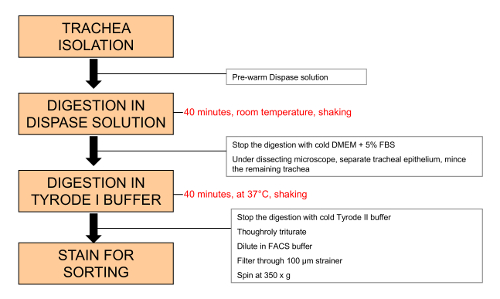
Figure 1: Schema of proposed steps for isolation of tracheal brush cells. The protocol includes 2 major steps: after dissection, the whole trachea is incubated in a high-dose dispase solution to separate the epithelium; this is followed by digestion of the epithelial sheet with papain in calcium containing Tyrode buffer (Tyrode I) and antibody staining. Please click here to view a larger version of this figure.
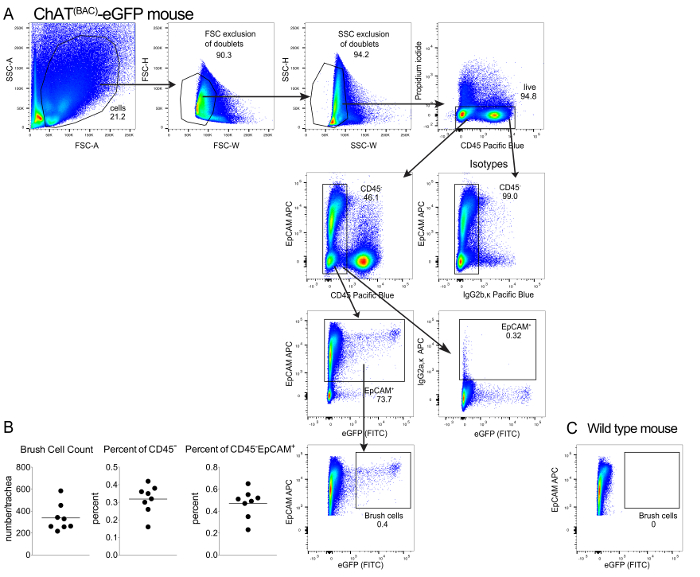
Figure 2: Representative flow cytometry analysis of tracheal brush cells. (A). Schematic representation of the flow cytometry gating strategy. Single cells were gated based on their forward and side scatter characteristics and live cells were chosen based on the exclusion of Propidium Iodide. CD45 low/negative cells were chosen based on the isotype control and evaluated for their expression of EpCAM. EpCAM positive cells were considered brush cells if they expressed eGFP fluorescent in the FITC channel. (B). Number of cholinergic tracheal brush cells per mouse trachea and frequency presented as percent of all CD45low/- cells and as a percent of CD45low/-EpCAM+ cells. Each dot represents a separate mouse. Data are from 3 separate experiments with 2-3 mice each. (C). Absence of eGFP positive brush cells in a tracheal epithelial digest from a wild type mouse stained for CD45 and EpCAM. Please click here to view a larger version of this figure.
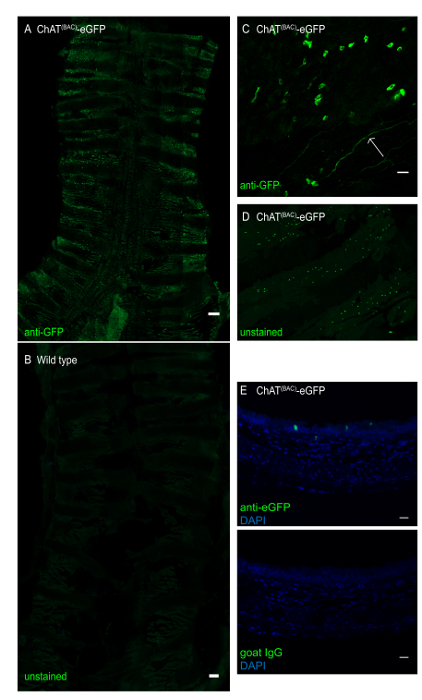
Figure 3: Whole mount of mouse trachea of a ChAT(BAC)-eGFP mouse. The trachea was opened longitudinally and stained with anti-GFP antibody to enhance the eGFP green fluorescence signal. (A) Whole tracheal mount of ChAT(BAC)-eGFP mouse, intensely green fluorescent cells represent brush cells; scale bar 1 mm; (B) whole tracheal mount of a wild type mouse demonstrating absence of brush cells; Scale bar = 1 mm; (C) magnification of the epithelial layer demonstrating irregularly shaped brush cells (green cells) as wells as a cholinergic nerve ending (arrow); (D) whole tracheal mount of a trachea of a ChAT(BAC)-eGFP imaged for GFP without antibody-enhancement of the fluorescence signal; (E) cross-section of a paraffin-embedded mouse trachea of a ChAT(BAC)-eGFP mouse stained with anti-GFP (green) or goat IgG control and DAPI (blue); Scale bars = 20 μm (C-E). Please click here to view a larger version of this figure.
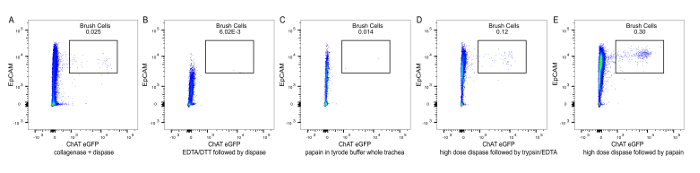
Figure 4: Comparison of different tracheal digestion protocols. (A) The trachea was minced and incubated with 650 U/mL collagenase IV and 1.84 U/mL dispase supplemented with DNase I for 45 min. This experiment was repeated 2 times. (B) The whole trachea was incubated with 2.5 mM EDTA and 0.75 mM DTT supplemented with DNase I for 20 min; the epithelial cells were dislodged and further digested with 0.01 U/mL dispase and DNase I for 10 min. This experiment was repeated 2 times. (C) The trachea was minced and incubated with 35 U/mL of papain in Tyrode I buffer for 45 min; residual papain activity was inhibited with leupeptin. This experiment was performed once. (D) The whole trachea was incubated with high dose dispase (16 U/mL) for 30 min to separate the epithelial sheet, followed by incubation of the epithelial sheet with 0.1% trypsin and 1.6 mM EDTA for 30 min. This experiment was repeated 2 times. (E). The whole trachea was incubated with high dose dispase (16 U/mL) for 30 min followed by digestion of the epithelial sheet with 26 U/mL papain in Tyrode buffer for 30 min. The residual papain activity was inhibited with leupeptin and high volume dilution. Please click here to view a larger version of this figure.
