A representative trace of mitochondrial oxygen consumption using the protocol described above is provided in Figure 2. The pyruvate and malate injected in the chambers along with the permeabilized muscle fibers are referred to the CI-LEAK respiration, i.e., when the complex I of the ETS is stimulated by the NADH produced through oxidation of pyruvate and malate via the tricarboxylic acid cycle (CI). During this respiration rate, the mitochondrial oxygen is mainly maintained to compensate for the proton leak, i.e., protons crossing from the intermembrane space to the mitochondrial matrix, when ATP synthase is not active (LEAK)30. When the ADP is added, the ATP synthase is activated and the electrons are transferred from the complex I to the complex IV with concurrent phosphorylation of ADP into ATP (CI-OXPHOS), resulting in increased mitochondrial oxygen consumption. The OXPHOS coupling ratio is calculated as CI-OXPHOS/CI-LEAK and is usually taken as a good indicator of mitochondrial quality and of mitochondrial coupling30. In Drosophila, this ratio should at least exceed 6.0 (Figure 3). If it is below this value, it could indicate a problem with the tissue preparation or a mitochondrial dysfunction. The addition of cytochrome c allows the determination of the integrity of the outer mitochondrial membrane and is therefore used as a quality control of the preparation (Figure 4). Indeed, cytochrome c is loosely bound to the inner mitochondrial membrane and is typically washed away if the outer mitochondrial membrane is damaged during the permeabilization process. As a result, adding exogenous cytochrome c will significantly increase oxygen consumption if the outer mitochondrial membrane is damaged and the endogenous cytochrome c is lost. An increase of less than 10-15% in oxygen consumption (Figure 4) usually illustrates appropriate integrity of the outer mitochondrial membrane29. In Figure 3 and 4, the green traces were obtained from non-adequate permeabilization and handling of the samples (excessive tearing of the tissue for Figure 3, and weighed after a longer time on the absorbent surface for Figure 4), whereas the red traces represent samples adequately permeabilized and handled. These results highlight that appropriate permeabilization conditions are crucial for reliable assessment of mitochondrial oxygen consumption.
The following substrates used during the experiments provide electrons to the ubiquinone and allowed to increase the electron flux into the ETS. Proline is an amino acid that can be used as energy substrate notably in insects23, but also in mammals during acute starvation or during pathological conditions37. The addition of proline in the chambers allows to evaluate the contribution of proline dehydrogenase (ProDH) to the electron flux in the ETS, when electrons are flowing from both complex I and ProDH and are participating to the OXPHOS process (CI+ProDH-OXPHOS). With addition of succinate, the contribution of complex II (succinate dehydrogenase) to the ETS can be observed (CI+ProDH+CII-OXPHOS). Injection of glycerol-3-phosphate (G3P), further increases the mitochondrial oxygen consumption as this substrate stimulates the mitochondrial glycerol-3-phosphate dehydrogenase (G3PDH) which is part of the glycerophosphate shuttle and transfer electrons to the ETS (CI+ProDH+CII+G3PDH-OXPHOS). Both ProDH and G3PDH have been shown to be particularly active in several species of insects and are therefore important in this protocol with Drosophila20,21,22,23,32.
When the uncoupler FCCP is added, the non-coupled respiration (ETS state) is obtained, i.e., the maximal oxygen consumption representing the maximal capacity of the ETS (CI+ProDH+CII+G3P-ETS). The FCCP is a protonophore that transports the protons from the intermembrane space to the mitochondrial matrix without passing through complex V. The FCCP has to be carefully titrated, as concentrations below optimum result in non-maximal oxygen consumption and non-stable respiration rates while concentrations above optimum result in inhibition of mitochondrial oxygen consumption (Figure 5). Once the substrates and the uncoupler are added, inhibition of complexes I, II, and III can be sequentially performed using rotenone, malonate, and antimycin A, respectively. With each inhibitor used, a decrease in mitochondrial oxygen consumption is observed, until reaching the lowest oxygen consumption after the addition of antimycin A. The rate reached after the addition of all the inhibitors is the residual oxygen consumption30. It represents the oxygen consumption due to non-mitochondrial oxidative reactions such as oxygenase reactions and reactive species production for example, and has therefore to be subtracted from all the other rates measured.
TMPD is an artificial electron transporter that provides electrons directly to complex IV, bypassing all the complexes upstream, allowing measurement of the complex IV maximal respiration capacity. This substrate is however prone to autoxidation, so ascorbate has to be added prior to TMPD to limit but not completely avoid this autoxidation. To correct for the remaining autoxidation of TMPD, the Complex IV inhibitor sodium azide is added to the chambers and the oxygen consumption is then recorded for 10-15 minutes. This oxygen consumption therefore represents the chemical background, or in other words the autoxidation of TMPD that should be taken into account for the calculation of complex IV maximal respiration capacity.
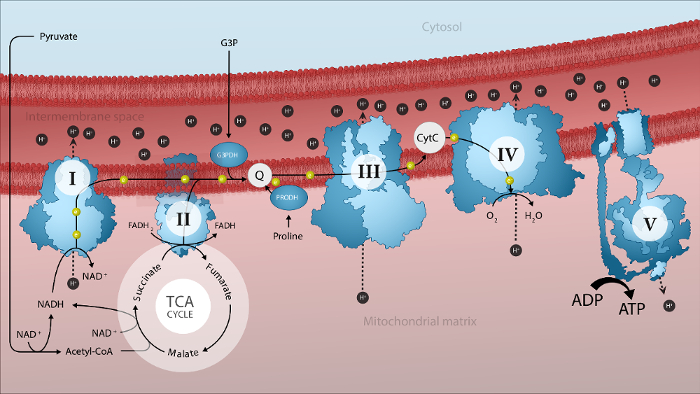
Figure 1. Schematic representation of mitochondrial electron transport by the electron transport system and oxidative phosphorylation in the mitochondrial inner membrane. I: complex I; II: complex II; III: complex III; IV: complex IV; V: ATP synthase; Acetyl-CoA: acetyl coenzyme A; Cyt c: cytochrome c; e–: electron; G3P: glycerol-3-phosphate; G3PDH: mitochondrial glycerol-3-phosphate dehydrogenase; H+: proton; PRODH: proline dehydrogenase; TCA: tricarboxylic acid cycle. Please click here to view a larger version of this figure.
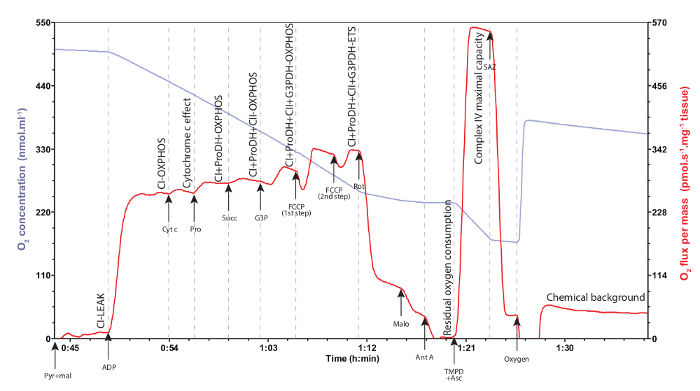
Figure 2: Representative traces of mitochondrial oxygen concentration (blue trace) and consumption (red trace) on permeabilized thoraxes of Drosophila. Each injection is represented with an arrow indicating the compound injected (Pyr: pyruvate; mal: malate; ADP; Cyt c: cytochrome c; Pro: proline; Succ: succinate; G3P: glycerol-3-phosphate; FCCP: carbonyl cyanide 4-(trifluoromethoxy)phenylhydrazone; Rot: rotenone; Malo: malonate; Ant A: antimycin A; TMPD: N,N,N',N,-Tetramethyl-p-phenylenediamine; Asc: ascorbate; SAZ: sodium azide). Each mitochondrial oxygen consumption rate is denoted on the graph, when the oxygen consumption (red trace) has been stabilized. Cytochrome c effect denotes the integrity of the outer mitochondrial membrane (see text for details). Residual oxygen consumption represents the oxygen consumption due to oxidative side reactions in the cells and has to be subtracted to all the other rates measured. Chemical background denotes the oxygen consumption only due to autoxidation of TMPD and has to be taken into account for the calculation of the complex IV maximal respiration capacity. Please click here to view a larger version of this figure.
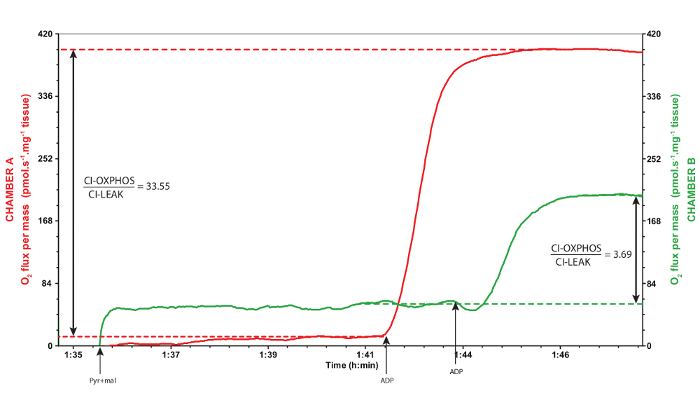
Figure 3: Representative traces of a valid (red trace, chamber A) and inadequate (green trace, chamber B) responses to the addition of ADP. The red trace corresponds to the chamber A and the green trace corresponds to the chamber B, during the same experiment. The red trace was obtained from thoraxes adequately permeabilized whereas the green trace was obtained after excessively tearing the tissues during the permeabilization. When ADP is added, an increase in the oxygen consumption is expected, like the red trace. OXPHOS coupling ratios are calculated as CI-OXPHOS/CI-LEAK and are presented in the graph. A ratio of less than 6.0 in Drosophila suggests a problem in the mitochondrial coupling and is characteristic of sample degradation or of mitochondrial dysfunction represented by the green trace. Pyr: pyruvate; mal: malate. Please click here to view a larger version of this figure.
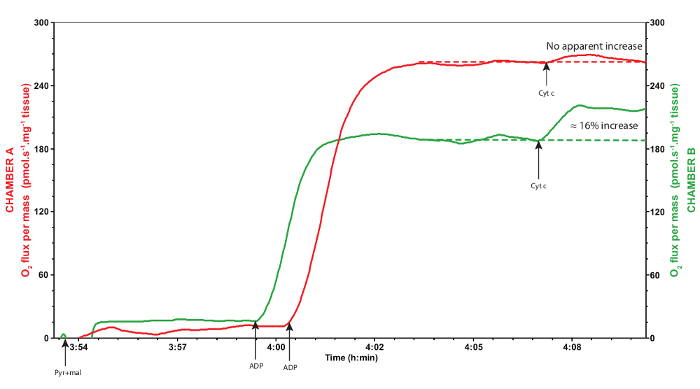
Figure 4: Representative traces of a valid (red trace, chamber A) and inadequate (green trace, chamber B) response to the addition of cytochrome c. The red trace corresponds to the chamber A and the green trace corresponds to the chamber B, during the same experiment. Red trace was obtained from thoraxes adequately permeabilized and handled, whereas the green trace was obtained after the thoraxes were purposely dried for a longer time on the absorbent surface before weighing. Cytochrome c does not usually increase mitochondrial oxygen consumption, as seen on the red trace, denoting that the outer mitochondrial membrane is intact. If, however, addition of exogenous cytochrome c significantly increases mitochondrial oxygen consumption of more than 15%, as seen on the green trace, it suggests that the outer mitochondrial membrane is damaged and hence that the sample is degraded and should be discarded. Pyr: pyruvate; mal: malate; Cyt c: cytochrome c. Please click here to view a larger version of this figure.
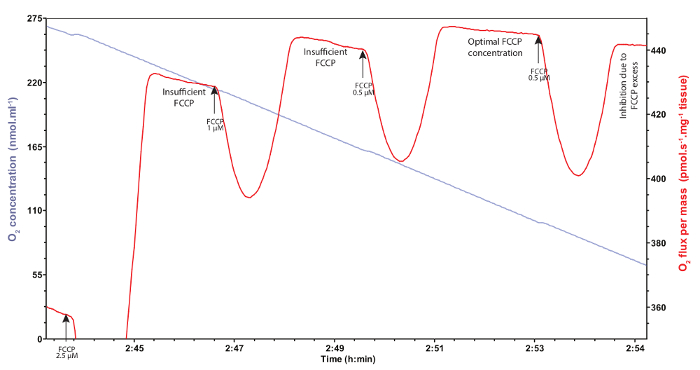
Figure 5: Representative traces of mitochondrial oxygen concentration (blue trace) and consumption (red trace) after titration of FCCP. Different injections of FCCP at several concentrations have to be carefully performed to determine the maximal oxygen consumption triggered by this uncoupler. Insufficient concentrations do not allow the assessment of maximal oxygen consumption and are characterized by non-stable respiration rates, whereas FCCP excess inhibits mitochondrial oxygen consumption. Arrows denote the different injections of FCCP. Please click here to view a larger version of this figure.
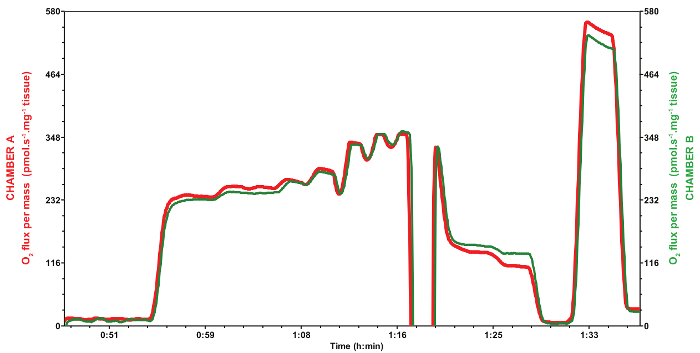
Figure 6: Reproducibility of mitochondrial oxygen consumption rates during a typical experiment obtained using the two different chambers (red trace, chamber A; green trace, chamber B) of a respirometer. The red trace corresponds to the chamber A and the green trace corresponds to the chamber B, during the same experiment using Drosophila from the same age, sex, strain and raised on the same diet, as well as the protocol described. Both samples were normalized with the mass of dry tissue weighed as described in the protocol (0.53 and 0.66 mg for chambers A and B, respectively). Please click here to view a larger version of this figure.
