1. Preparation of Buffers and Solutions
- To prepare 1 L of phosphate buffered saline (PBS, pH 7), dissolve 8 g of NaCl, 0.2 g of KCl, 1.44 g of Na2HPO4, 0.24 g of KH2PO4 in 800 mL of distillated water. Adjust the pH to 7.0 with HCl, and then add water for a final volume of 1 L. PBS can be made as a 10x stock solution and stored at room temperature.
- To prepare 10 mL of fixing solution, add 1 mL of 37% formaldehyde to 9 mL of PBS to reach a final concentration of 3.7%. Prepare this solution just before use.
Caution: Use gloves to handle the formaldehyde solution. - To prepare the quenching-solution (500 mM), dissolve 8.3 g of KI in 100 mL of distilled water. Prepare this solution before use and store it at room temperature.
- To prepare BODIPY 10 mM stock solution, dissolve 10 mg of BODIPY 493/503 in 3.8 mL of dimethyl sulfoxide (DMSO). Keep 100 µL aliquots in the dark at -70 °C.
- To prepare BODIPY 5 µM-quenching solution, add 1 µL of 10 mM BODIPY 493/503 to 2 mL of 500 mM quenching solution. For the LFR assay, a final concentration of 5 µM of BODIPY is required.
- To prepare the mineral solution, add 0.06 g of H3BO3, 0.14 g of MnCl2-4H2O, 0.4 g of ZnCl2, 0.04 g of Na2MoO4-2H2O, 0.1 g of FeCl3-6H2O and 0.4 g of CuSO4-5H2O to 1 L of distilled water.
- To prepare the salt solution, add 16 g of KH2PO4, 4 g of Na2SO4, 8 g of KCl, 2 g of MgSO4, 1 g of CaCl2, and 8 mL of mineral solution to 1 L of distilled water.
- To prepare the minimal medium without a nitrogen source for U. maydis FB2 (a2b2), add 10 g of glucose and 62.5 mL of salt solution to 1 L of distilled water, according to Holliday, et al.31. Adjust the pH to 7.0 and then dispense 100 mL of the media in 250 mL flasks and sterilize them. Autoclave for 20 min at 15 psi (1.05 kg/cm2) on liquid cycle or filter sterilize.
- To prepare the yeast peptone dextrose medium (YPD), add 5 g of yeast extract, 2.5 g of peptone, and 5 g of glucose to 1 L of distilled water. Dispense 100 mL of the solution in 250 mL flasks and sterilize them. Autoclave for 20 min at 15 psi (1.05 kg/cm2) on liquid cycle or filter sterilize.
2. Culture Condition and Cell Fixation
NOTE: Preserve the structure of the LDs by fixing the cells as soon as possible. The fixation can be done in microcentrifuge tubes. Fixed cells can be kept at 4 °C for up to one month if dehydration is avoided leaving a thin layer of water.
- Grow U. maydis cells under culture conditions relevant for the study. Start the culture with an initial OD600 of 0.05 (1.15 × 106 cells/ mL). Incubate the cells at 28 °C, 180 rpm for 24 h. An OD600 of 1 corresponds to 2.3 × 107 cells/ mL.
- At the selected times, withdraw an aliquot of the cells. For U. maydis, withdraw 5 mL every 2 h during the first 8 h. After 8 h of growth, aliquots of 2 mL are enough.
- Measure the optical density at 600 nm of each aliquot.
- Centrifuge the cell suspension at 14,000 x g for 1 min at 4 °C using a tabletop centrifuge. Discard the supernatant.
- Suspend the pellet of the cells in 1 mL of PBS-formaldehyde 3.7% buffer. Incubate the cells for 15 min at room temperature.
- After the incubation, centrifuge the cell suspension at 14,000 x g for 1 min at 4 °C. Discard the supernatant.
- Wash the cells pellet twice with a similar volume of distilled water (1 mL). Centrifuge the cell suspension at 14,000 x g for 1 min at 4 °C. Discard the supernatant after each washing step.
- Adjust the cell pellet to 5 OD600 (1.15 x 108 cell/ mL) with distilled water (Equation 1).
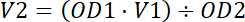 Equation 1.
Equation 1. - Keep the sample at 4 °C until its use.
3. Liquid Fluorescence Recovery Assay (LFR)
- Turn on the spectrophotometer and open the Skanlt software. Click on NEW SESSION, and choose START and then Varioskan Lux. Select PROTOCOL from the session tree, enter the settings for the fluorescence wavelengths (excitation 485 nm/emission 510 nm; optical density 600 nm) and choose automatic photomultiplier gain. For excitation bandwidth, select 12 nm. In the optics, select top (excitation of the sample is from the top of the well).
- Enter a gentle and continuous agitation (300 rpm). Select RUN PLATE OUT, and PAUSE UNTIL THE USER ACTION (time used for the addition of the sample to the wells). Repeat the protocol five times. Click SAVE and write a name for the session on the SESSION NAME FIELD. The protocol is now ready for the samples analyses.
- Add 200 µL of the BODIPY 5 µM- quenched solution to each well of a 96 well black-clear bottom plate.
- Place the plate inside the spectrophotometer chamber and incubate 5 min at 30 °C. From this point, protect the plate from light as much as possible.
- Click the START button and read the fluorescence (excitation 485/emission 510) and the OD600 corresponding to the blanks.
- Add 5 µL of the formaldehyde-fixed cell suspension to the wells during the pause time. Mix the samples carefully with the pipette. Put the plate inside the spectrophotometer and click CONTINUE. Then, the samples will be processed according to the protocol designed above.
- Repeat three successive additions of 5 µL of cell suspension. Make sure that the cells do not precipitate. (Figure 1).
4. Calculations of The LD Index
- For each sample in the microplate, plot a graph of fluorescence and absorbance against the volume of each successive addition (5 points including the blank). In this study, use Microsoft Excel software for the calculations. To plot the data, enter the volume, fluorescence, and absorbance data in the first, second and third column of the datasheet, respectively. Then, select the whole data with the cursor, click INSERT and select (XY) SCATTER.
- Select the points for each line in the graph and insert the respective TREND LINE with the SHOW EQUATION box selected. Write down the values of the slopes of the two straight lines, according to equation 2:
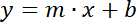 Equation 2
Equation 2
Where y= fluorescence or optical density, x = volume of sample (0, 5, 10, 15, 20 µL), m = slope of the fluorescence or optical density line, and b = y-intercept. - Divide the slope of the fluorescence line by the slope of the optical density line to get the LD index, equation 3.
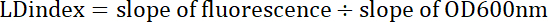 Equation 3
Equation 3
The LD index corresponds to the quotient of fluorescence and optical density slopes.
5. Data Quality Analysis
- Calculate the correlation coefficient of each straight line: click on the FUNCTION WIZARD (fx) and choose CORREL. If r <0.9, discard the data and repeat the readings. If r ≥ 0.9, readings are reliable.
LFR, with BODIPY 493/503 as the fluorescent probe is a reliable and easy method to study the dynamic accumulation of LDs in U. maydis, regardless the growth conditions. When cells were cultured in YPD-medium, there was an increase in the LD index during the exponential phase, followed by a decline in the stationary phase (Figure 2A). In contrast, in cells grown under nitrogen starvation, there was a steady increase in LD index in both the exponential and stationary phases (Figure 2B). Because both cultures were started with the same optical density (similar number of cells), the results show the high capacity of the LFR assay to detect small changes in the lipid content (Figure 2). The sensitivity of the method was corroborated by confocal microscopy: YPD-cells contain many small LDs throughout the whole cell, while under nitrogen starvation there was an important production of large LDs, typically 4-5 LDs per cells, (Figure 2, right panels A and B, respectively). Since the LD index is not an absolute value of the neutral lipid content, a standard curve relating the LD index with the amount of triacylglycerols should be constructed. This approach was used to study the dynamics of TAGs synthesis in S. cerevisiae20. Based on each standard curve, the LFR assay can be used to determinate the rate of lipid accumulation in cells.
The LD index can be followed in the presence of soraphen A, a specific inhibitor of the acetyl-CoA carboxylase, an enzyme that is essential for the synthesis of fatty acids contained in the TAGs stored in LDs25. The inhibitor was added to the culture medium after 2 h of growth in the absence of a nitrogen source, a condition in which cells showed the higher rate of lipid accumulation (Figure 2B). In accordance with previous reports in S. cerevisiae, the addition of soraphen-A resulted in the full inhibition of LD accumulation (Figure 2B, light blue) 20,21.
To further investigate the mobilization of LDs in U. maydis, cells grown under nitrogen starvation for 72 h were transferred to YPD-medium (Figure 3). LD index decreased following an exponential function. After 24 h, the LD index reached similar values to that observed in cells cultured in YPD-medium without the stress of nitrogen absence (Figure 2A). Since the cells were able to mobilize the LDs accumulated during nitrogen starvation, the result suggests a reduction in the TAGs content. A similar response was reported for S. cerevisiae during the study of phosphatases involved in the regulation of lipid metabolism21.
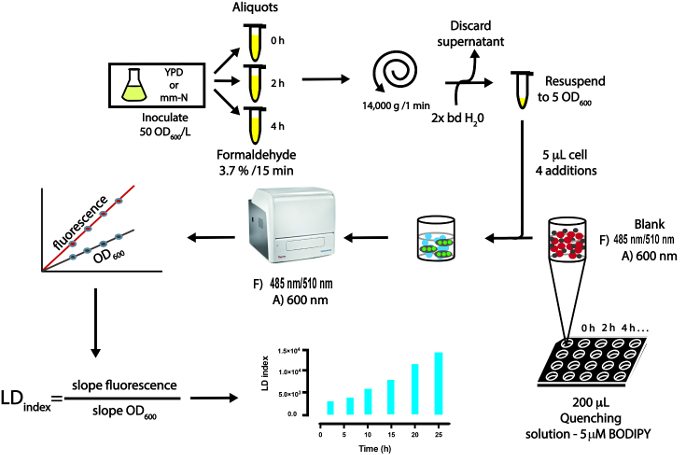
Figure 1: Schematic representation of the steps involved in the LFR. The steps of culture conditions are representative for U. maydis yeast but can be adapted to other microorganism or type of cells. URF, Units of Recovery Fluorescence; bd, bidistilled water. Please click here to view a larger version of this figure.
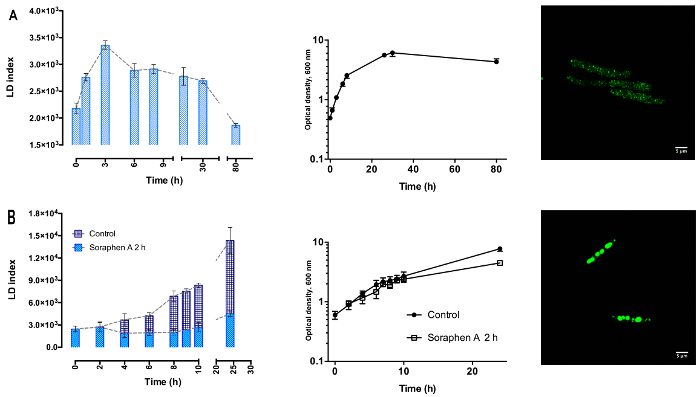
Figure 2: Dynamics of lipid droplets accumulation in U. maydis. Yeast cells were grown for 24 h in YPD-medium, harvested and suspended in (A) fresh YPD-medium (Top left panel) or in (B) minimal medium without a nitrogen source (bottom left panel-dark blue). Under nitrogen starvation, the increase in LD index was inhibited by adding 192 nM soraphen A in the culture media after 2 h of growth (B, light blue). n=3, data are the mean ± SEM. Right panel: confocal microscopy of LDs in cell harvested at 24 h. (A) Top panel: YPD- cells. (B) Bottom panel: cells without a nitrogen source. Please click here to view a larger version of this figure.
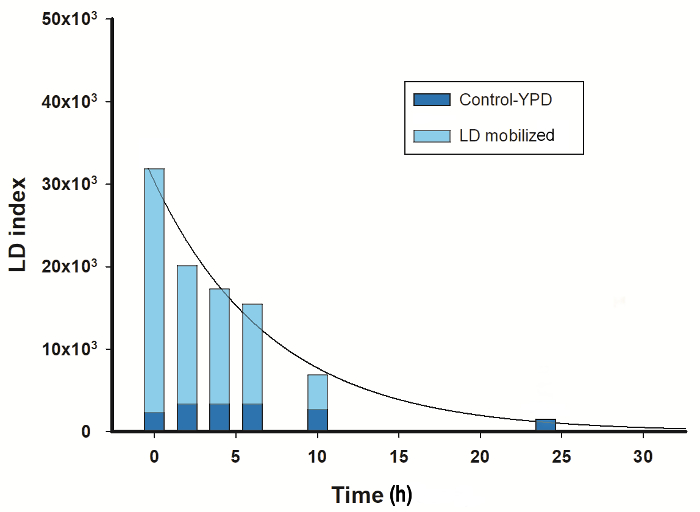
Figure 3: LDs mobilization. Yeast cells were grown for 24 h in YPD-medium, harvested and grown in fresh YPD for 24 h (control) or in minimal medium without a nitrogen source for 72 h, then transferred to fresh-YPD medium and incubated for additional 24 h. Aliquots were withdrawn at the indicated times for both cultures to measure the mobilization of LDs by LFR. Data was fitted by an exponential decay equation (y = A·e-kt). Please click here to view a larger version of this figure.
