In this protocol, three-slit grids were used for picking up serial sections. The grids are made of nickel or copper. The serial sections are placed on the middle slit. The slits on both sides are necessary to view the sections when picking them up with the grid. To keep the grids parallel with the serial sections when picking them up with tweezers (Figure 11d), the handle is bent (Figure 6c, right). A small handle is advantageous to prevent bending of the main part of the grid, which causes serious problems in picking up sections, and to prevent dropping the sections during staining. This grid has advantages over the conventional 2.0 mm x 1.0 mm single-hole grid. Namely, sections are supported directly by two metal bars 0.4 mm apart (narrower than 1.0 mm) and by the Formvar support film in a three-slit grid, while sections are supported only by the support film in a conventional single-hole grid. Specimen drift can therefore be prevented in photographing at high magnification using a three-slit grid.
Figure 15 shows a low magnification view of five grids that carry serial sections. Since each section sticks together, it is easy to find the same cell in the next section even at higher magnification. Figure 16 and 17 are examples of serial images of cell sections. Because sections are securely supported on 3-slit grids, photos at high magnification (x 50,000) are easily taken without specimen drift. Fibers of DNA 2 nm in diameter were photographed in these studies9, 12.
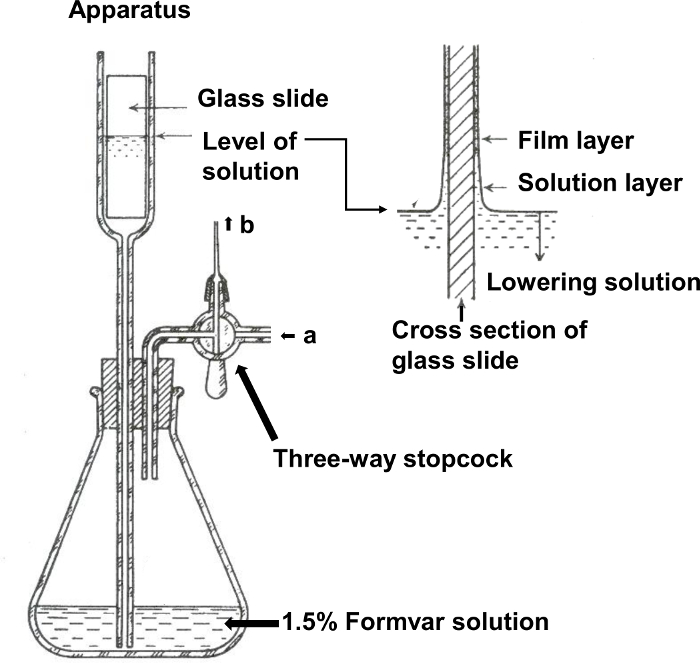
Figure 1. Formvar support film-making apparatus. One-half of the glass slide is dipped in Formvar solution in the upper column of the apparatus by pressing the solution with air through 'a' using a rubber ball. The solution is drained from the column by opening the three-way stopcock and releasing air through 'b' to reduce pressure. (Reproduced from Yamaguchi and Adachi, 201119 with permission). Please click here to view a larger version of this figure.
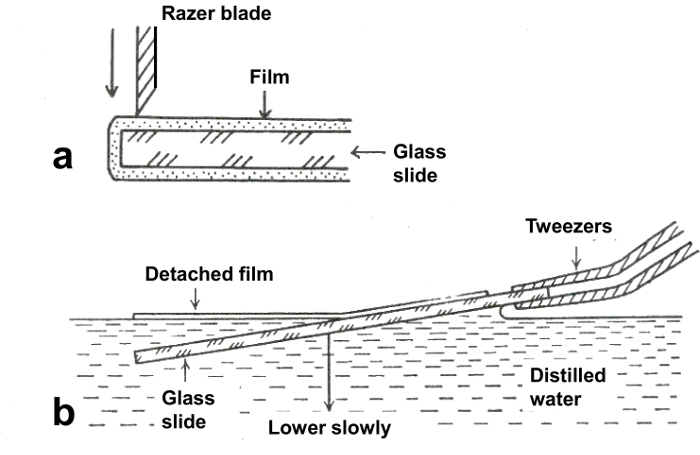
Figure 2. Method for releasing the film from the glass slide on water. (a) The four edges of the film on the glass slide are scraped off using razor blade. (b) Formvar film is floated off on the water by lowering the glass slide slowly at a low angle. (Reproduced from Yamaguchi and Adachi, 201119 with permission). Please click here to view a larger version of this figure.

Figure 3. Formvar support film on an aluminum rack. (a) The Formvar film is scooped up from the water with an aluminum rack. (b) The rack is kept in the desiccator until use. Please click here to view a larger version of this figure.

Figure 4. Trimming of the specimen block with an ultrasonic trimming blade and a razor blade under a stereomicroscope. (a) The presence of the specimen is confirmed by observing the specimen block under a light microscope. The block shown in the figure contained microorganisms associated with a scale-worm chaeta (about 1.7 mm in length) collected from the deep sea12, 14. (b) Trimming stage. (c) Whole image of a stereomicroscope. Please click here to view a larger version of this figure.
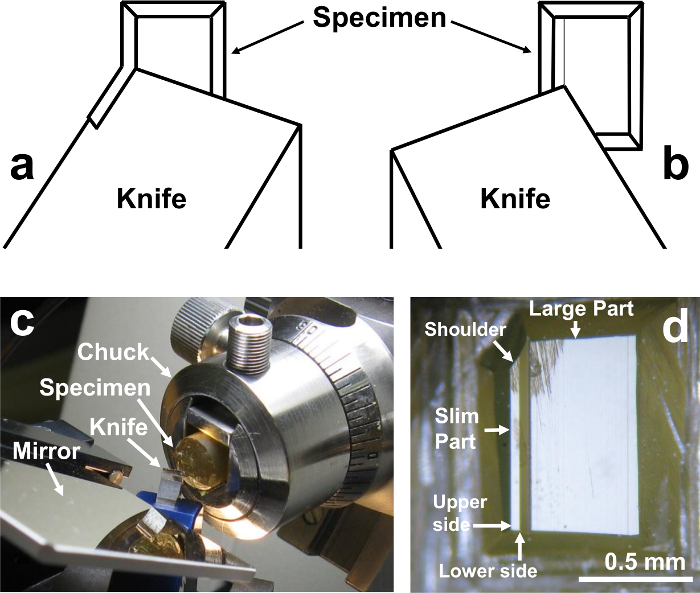
Figure 5. Trimming of the specimen block with diamond knife using microtome. (a) Illustration for cutting the left edge. (b) The block face is cut at a position of about 100 nm from the left edge of the specimen. (c) A mirror (Mesa cut, M) placed on the knife stage. (d) Specimen surface. Note that the upper side and the lower side of the slim part are smooth and perfectly parallel. Also note that the shoulder is cut to mark the direction of cutting (see Figure 8). Please click here to view a larger version of this figure.

Figure 6. Neoprene treatment on the specimen surface. (a) In order to place the specimen chuck back at exactly the original position, the chuck and the chuck holder of the microtome are marked with a tape and cut at the boundary. (b) Wetting the specimen with neoprene solution using Pasteur pipette. (c) Three-slit grids for picking up serial sections. The handle is bent to 60 degrees (c, right). Please click here to view a larger version of this figure.

Figure 7. Plastic cover of the microtome and wooden armrest. The plastic cover is useful to prevent airflow during ultrathin sectioning. The wooden armrest is used for doing delicate work such as retrieving serial sections. Please click here to view a larger version of this figure.
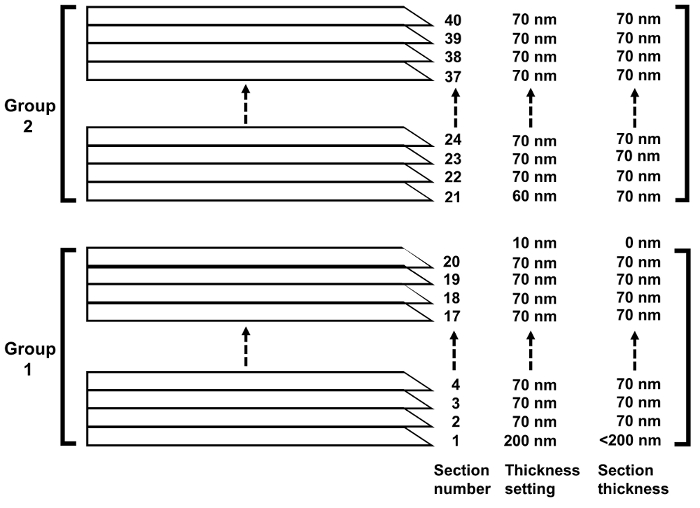
Figure 8. Setting section thickness and obtaining sections at the proper thickness. After the number of serial sections has reached 20, section thickness is set to 10 nm. Since the microtome cannot cut 10 nm-thick sections, no new section appears, and the previously cut sections become separated from the knife edge. Please click here to view a larger version of this figure.

Figure 9. Example of ultrathin sections obtained through the protocol. Note that the five groups of serial sections of about 1.8 mm long are already separated after cutting. In this particular case, the color of the center of the ultrathin sections is yellowish due to the presence of the specimen. The other parts of the section, which do not contain the specimen and consist of resin only, are silver in color. Please click here to view a larger version of this figure.

Figure 10. 'Section-holding loop'. (a) Top view (upper part) and bottom view (lower part of photo) of the loop. (b) The Section-holding loop is used to hold the third group of serial sections. Please click here to view a larger version of this figure.
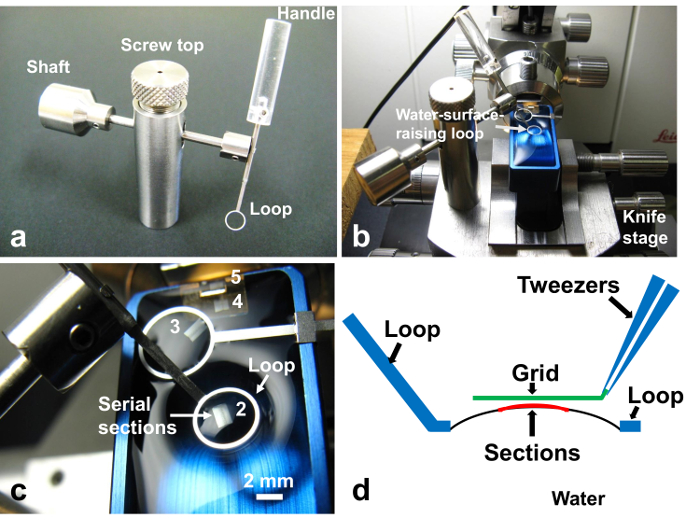
Figure 11. Retrieval of serial sections. (a) 'Water-surface-raising loop' (WSRL). The handle of the loop can be removed from the shaft, which is movable. The loop can also be moved up and down by turning the screw top. (b) The WSRL is fixed on the knife stage by a magnet. (c) Closer view of (b). (d) Diagram of the loop, water surface, sections, and grid during retrieval (side view). Ultrathin sections can be retrieved precisely to the slit grid by lowering the bare grid parallel to the ultrathin sections. Please click here to view a larger version of this figure.

Figure 12. Placing the grid on Formvar film. The grid holding the sections is placed along with a tiny drop of water on the Formvar film on aluminum rack. Please click here to view a larger version of this figure.

Figure 13. Staining tube16. A groove 0.6 mm deep is made along the long axis using a razor blade. Grids are placed in the groove in the proper order. One end of the tube is cut (arrow head) to mark its direction. Please click here to view a larger version of this figure.

Figure 14. Placing the grids in the specimen holder. (a) The grids are placed in the specimen holder in the proper order. (b) The grids are then fixed with 'grid-fixers'. Please click here to view a larger version of this figure.

Figure 15. Serial sections mounted on slit grids. The figure shows 18 to 25 sections mounted on one grid. The numbers in each slit indicate the sequence number beginning from the first section. The first section is thicker than the others (see Figure 8). Note that the sections are attached together and have the same thickness. Please click here to view a larger version of this figure.
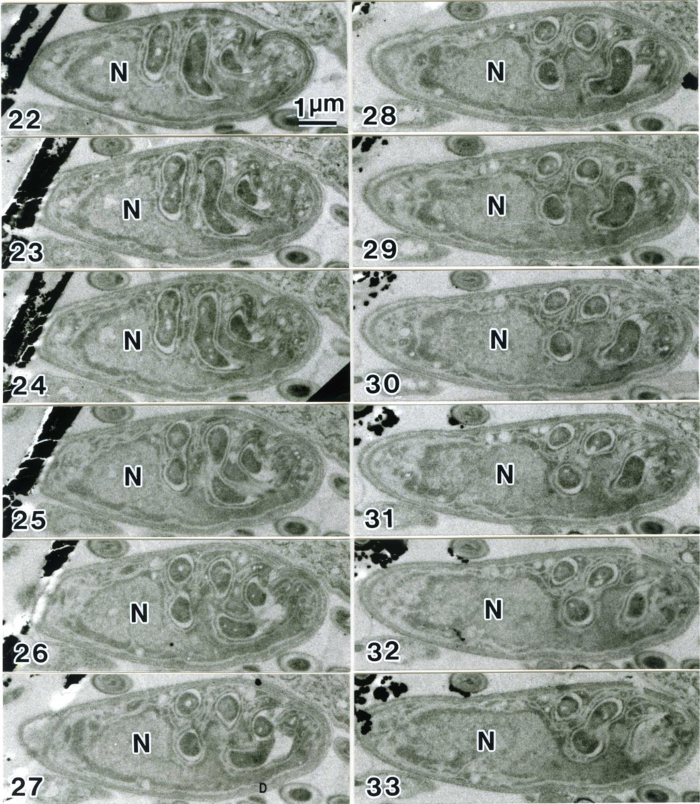
Figure 16. Serial sections of Parakaryon myojinensis12. The numbers at the lower left indicate the sequence relative to the first section. The figure shows 12 out of 67 complete sections. This cell was found to have a large nucleoid consisting of naked DNA fibers, with a single nucleoid membrane, and endosymbionts that resemble bacteria, but no mitochondria. Thus, this organism appears to be an intermediate life form evolving from prokaryote to eukaryote12. N, nucleoid. Reproduced from Yamaguchi and Worman, 2014.14 with permission. Please click here to view a larger version of this figure.
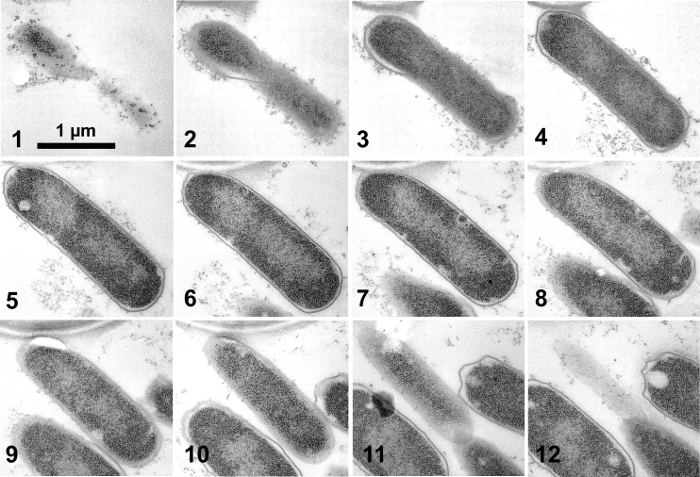
Figure 17. Serial sections of Escherichia coli9. The numbers at the lower left indicate the sequence relative to the first section. The figure shows 12 complete sections. This cell was found to contain 21,700 ribosomes9. Please click here to view a larger version of this figure.
