We used the above protocols to visualize steroidogenic organs and their interactive organs in D. melanogaster larvae and adult females. The overall scheme of protocols is shown in Figure 1.
The RG, including the PG (Figure 2D), is smaller and more transparent than the brain and is located at the anterior-dorsal side of the brain (Figure 2A-C and 3A-E). To label the PG cells, several groups have generated various types of antibodies against ecdysteroidogenic enzymes (i.e., Neverland23, Spookier24, Shroud20, Phantom25, Disembodied25, and Shadow26). Among them, anti-Shroud (Sro) antibody is reliably used to label the PG by immunostaining (Figure 2C, 2E, and 3C). Alternatively, the binary gene expression system, GAL4/UAS system27, can be used to express fluorescent protein genes in PG cells. Through the promoter analysis of the ecdysteroidogenic gene phantom (phm), the PG-specific promoter can induce gene expression exclusively in the PG28. Therefore, the PG cells can be specifically visualized by expressing GFP or RFP under the control of phm-GAL4#22 (Figure 3D).
To visualize the neuronal connection between the PG and the brain, a group of neurons can be labeled with mCD8::GFP under the control of various GAL4 drivers and antibodies (Figure 2C, 2E, 3D, and 3E). The FlyLight database of GAL4 line collections is optimized for labeling neurons29. Among them, GMR45C06-GAL4 labels PG-projecting neurons (Figure 2C and E). The immunostaining of GFP-expressing larvae with anti-GFP antibody is effective in enhancing GFP signals. Furthermore, TRH-GAL4>mCD8::GFP larvae show the stomatogastric nervous system, in which serotonergic neurons project to not only the PG, but also to the proventriculus (PV, insect foregut) and the pharyngeal muscles (PM) (Figure 3D and E)20,30. Therefore, it is critical to maintain the position of the RG, the brain, and the other surrounding tissues during fillet dissections (Figure 3).
In adult females, ovarian ecdysteroid affects many aspects of oogenesis, such as GSC proliferation, cyst differentiation, egg chamber growth, and stress response11. As in the PG, ecdysteroidogenic enzymes in the ovary are also visualized with the specific antibodies described above12,31. As a downstream event upon which ecdysteroids act, the number of GSCs in the germarium is our focus (Figure 4). Although the germarium is an assembly of multiple cell types, GSCs are specified by immunostaining with two GSC marker antibodies, 1B1 and DE-cadherin32. To visualize the innervation of the ovary, the ovary is dissected along with the brain, VNC, gut, crop, uterus, and spermatheca (Figure 5). Neurons are visualized by mCD8::GFP under the control of nSyb-GAL4, a pan-neuronal driver (Figure 5B and D). Muscles around the ovary, uterus, and gut are stained with dye-conjugated phalloidin.
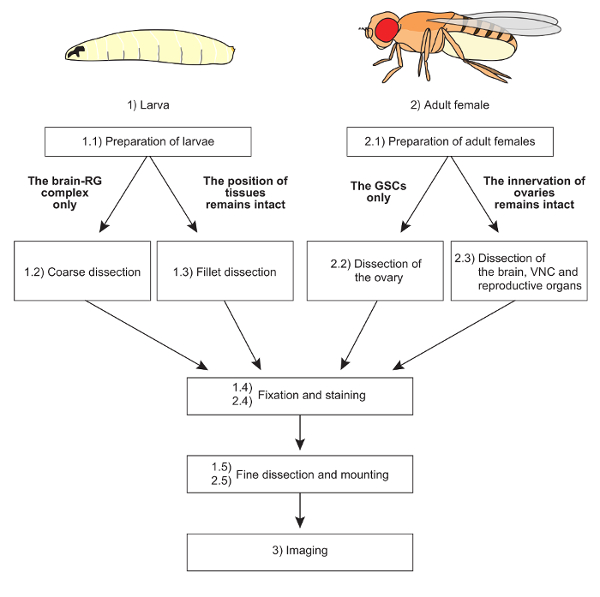
Figure 1: The overall scheme of protocols. Two distinct dissection methods are applicable to larvae and adult females, depending on the purpose of the experiments. The mounting methods are also devised according to sample conditions. Fixation, staining, and imaging techniques are basically common to all samples. Please click here to view a larger version of this figure.
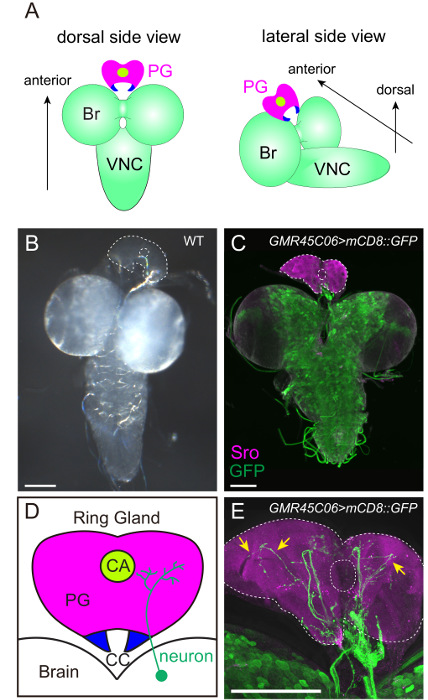
Figure 2: Visualization of the PG and the PG-projecting Neurons. (A and B) The brain-ring gland (RG) complex of the wild-type 3rd instar larva. In (B), the PG is outlined by dotted lines. The filamentous structure between the brain and the RG is the trachea. (C–E) The innervation of the PG was visualized with mCD8::GFP driven by GMR45C06-GAL4 in FlyLight collection. The PG and GFP-positive neurons were labeled with anti-Sro antibody (magenta) and anti-GFP antibody (green), respectively. The fluorescent image is merged with the transmitted-light image to specify the outline of tissues. In (D), the PG, the corpora allata (CA) and the corpora cardiaca (CC) are illustrated. The PG-projecting neurons are more prominent in the high-power image with 40X objective lens (arrows in E) than the low-power image with 10X objective lens (C). Scale bar = 100 µm. Please click here to view a larger version of this figure.
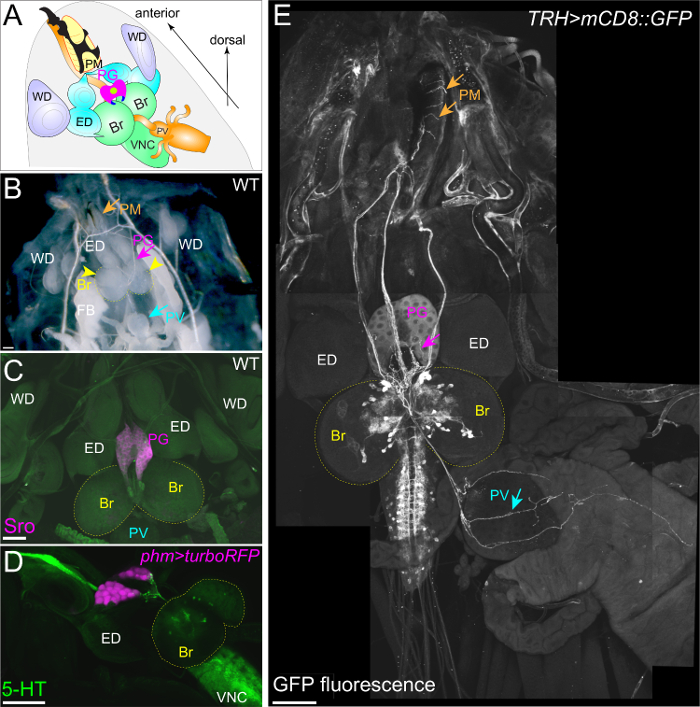
Figure 3: The Stomatogastric Nervous System Projects to the PG, the PV, and the PM. (A and B) The fillet dissection of a wild-type 3rd instar larva. The positions of the PG, brain (Br), ventral nerve cord (VNC), eye discs (ED), wing discs (WD), pharyngeal muscle (PM), and proventriculus (PV) remain intact. (C) The PG was exclusively labeled with anti-Sro antibody (magenta). (D) The PG was labeled with turboRFP driven by phm-GAL4#22. Serotonergic neurons were labeled with anti-5HT antibody (green). (E) The stomatogastric nervous system was visualized with mCD8::GFP driven by TRH-GAL4. Serotonergic neurons project to the PG, the PM, and the PV (arrows). The Scale bar = 100 µm. Please click here to view a larger version of this figure.
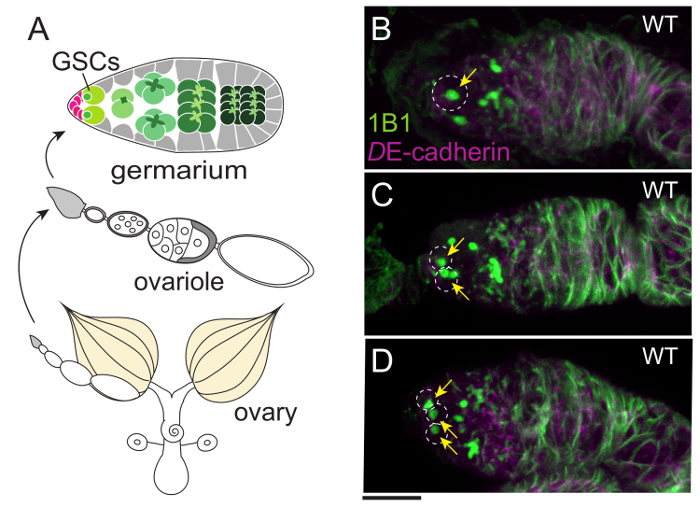
Figure 4: The Germaria of Wild-type Flies Containing One, Two, or Three GSCs. (A) An overview of the female reproductive organ. An ovary is composed of 16-20 ovarioles that are strings of egg chambers. The germarium, where GSCs are located, is at the tip of each ovariole. (B–D) One, two, or three GSCs are located in each germarium (white dotted circles) of the wild-type females. GSCs are stained with 1B1 antibody (green), which labels a spherical structure called the spectrosome (arrows) and a membranous cytoskeletal structure known as the fusome. The cell boundaries are visualized with anti-DE-cadherin antibody (magenta). The Scale bar = 10 µm. Please click here to view a larger version of this figure.
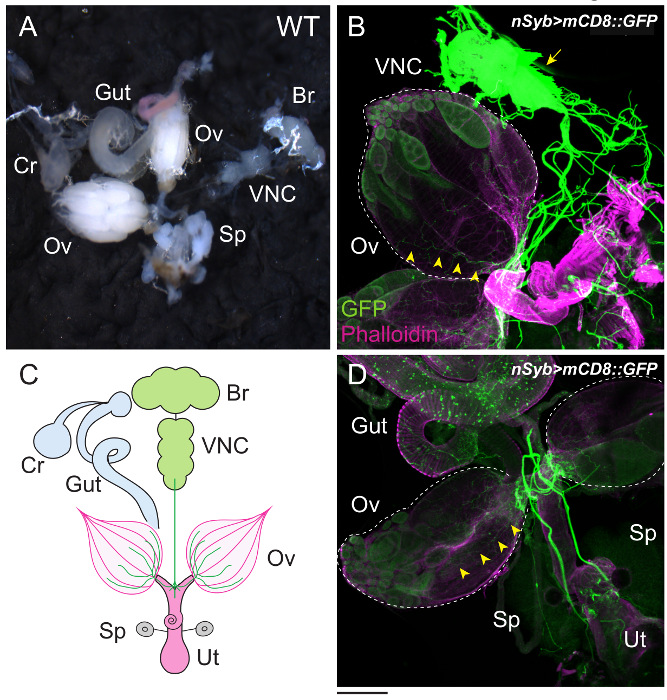
Figure 5: The Female Reproductive Organs and their Interactive Organs. (A) The ovary (Ov), gut (Gut), crop (Cr), brain (Br), ventral nerve cord (VNC), and spermatheca (Sp) were dissected under the microscope. (B–D) The innervation of the ovary was visualized with mCD8::GFP driven by nSyb-GAL4. GFP-positive neurons extend from the VNC (arrow), projecting to the surface of the ovaries (arrowheads). The ovaries are outlined by dotted lines. The samples were stained with anti-GFP antibody (green) and dye-conjugated phalloidin (magenta). Phalloidin associates with the filamentous actin of muscles around the ovaries (Ov) and uterus (Ut). The illustration is shown in C. The Scale bar = 200 µm. Please click here to view a larger version of this figure.
