The analysis of real-time raw data is described in Figure 1. First, platelets adhere to the reactive surface, resulting in a steady increase in recorded green fluorescence (Figure 1A, i), called adhesion. During this phase, there is little violet fluorescence, indicating that fibrin is not or is only marginally formed (Figure 1B). Upon initiation of coagulation, violet-fluorescing fibrin deposits rapidly (iii), and during that time, platelet green fluorescence increases at about the same rate, designated here as platelet accumulation (ii). A single experiment thus returns three rates of fluorescence increase (i, ii, and iii) as a surrogate marker for the velocity at which platelet and fibrin deposition takes place in this model. Furthermore, a moment of coagulation onset (moment-of-onset (iv)) is extrapolated, which is a determinant of platelet procoagulant potential.
In the absence of TF, coagulation initiation is slow and primarily runs through the contact pathway where the intrinsic tenase complex activates FX via direct activation of FIX6 by FXI and/or FXII. To demonstrate FXII dependence, 4 µM of corn trypsin inhibitor (CTI) was added to inhibit activated FXII (FXIIa)7. This inhibition did not affect platelet adhesion (Figure 2A), but coagulation did not start for the arbitrarily defined total duration of the perfusion experiment (Figure 2B and Supplementary Video 1).
In vitro, the contact pathway can be initiated by foreign materials like glass, or in clinical assays using a mineral material like kaolin. In vivo, activated platelets provide the negative charge8 through the membrane exposure of acidic phospholipids9, like phosphatidylserine, and/or through the release of polyphosphates (polyP)10,11. Our microfluidic real-time assay mimics the latter because, in a range of platelet concentrations, the hemostatic reaction depended on the platelet count (Figure 3). By increasing the number of platelets in the reconstituted sample, the rate of adhesion (Figure 3A), accumulation (Figure 3B, green), and coagulation (Figure 3B, violet) increased linearly. The moment-of-onset significantly shortened (Figure 3C) by increasing platelet concentration, suggesting that a threshold number of (activated) deposited platelets is required to trigger coagulation.
Upon tissue damage in vivo, however, TF-bearing cells will initiate the clotting of blood via the FVIIa-TF extrinsic tenase complex in the presence of Ca2+. This is mimicked in our experimental setup by post-coating the collagen-containing perfusion chambers with lipidated rhTF. In a paired analysis of channels coated with collagen only or in combination with rhTF, coagulation onset was significantly faster (Figure 4A). The rate of platelet adhesion was not linear, so no linear regression could be performed (data not shown). Both the rate of coagulation and platelet accumulation were not different between conditions (Figure 4B-4C, Supplementary Video 2).
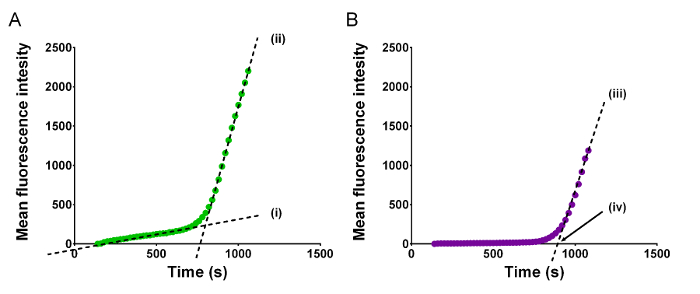
Figure 1: Regression analysis of platelet adhesion and coagulation in microfluidic perfusion chambers. This figure clarifies the parameters derived from real-time fluorescence raw data acquired during recalcified blood flow over a reactive surface. (A) Mean intensity of green fluorescence shows platelet deposition in function of perfusion time. The curve describes a bimodal process beginning with (i) slowly increasing linear platelet adhesion followed by (ii) rapidly growing linear accumulation. Both linear parts of the curve are regressed and the slope of these describes the two rates of thrombus formation by platelet fluorescence; (i) adhesion and (ii) accumulation. (B) Mean intensity of violet fluorescence shows deposition of fibrin in function of time. During platelet adhesion, violet fluorescence is essentially absent, while it quickly develops following initiation of coagulation. The (iv) moment-of-onset is defined as the intercept with the x-axis of the extrapolated linear regression of (iii) the second phase of thrombus formation by fibrin fluorescence designated here as coagulation. Please click here to view a larger version of this figure.
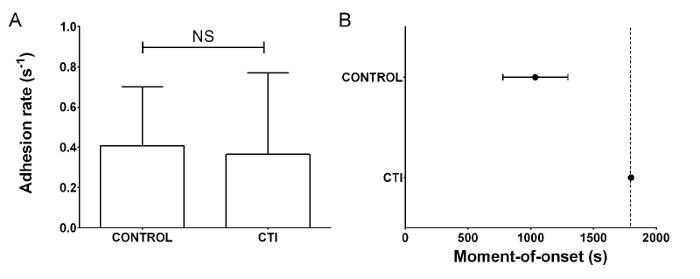
Figure 2: In the absence of TF, coagulation in collagen-coated perfusion flow chambers depends on FXIIa. Microfluidic perfusion of reconstituted blood containing CTI or control buffer (CONTROL) was performed at a shear rate of 1,000 s-1 for a total of 30 min. (A) The rate of platelet adhesion (s-1) was not dependent on CTI. (B) The moment-of-onset for CTI-treated samples was beyond the 30 min limit of the experiment (shown as a dotted line), demonstrating the dependence on FXIIa of this parameter. Bars and dots are mean values, whiskers are standard deviation. Statistical analysis was by paired t-test. (NS = not significant, n ≥3). Please click here to view a larger version of this figure.
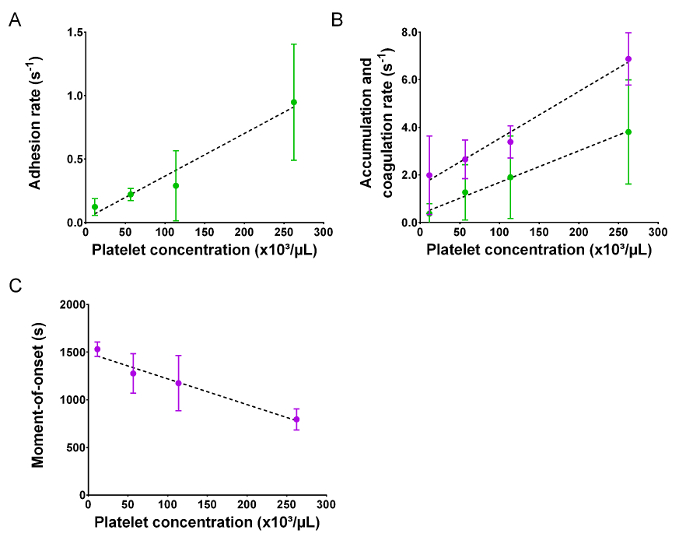
Figure 3: Coagulation under flow depends on the platelet number. Microfluidic perfusion experiments were performed in collagen-coated chambers with reconstituted blood in the presence of Ca2+ and varying platelet concentrations (n ≥3). (A) Rate of platelet adhesion (B) rate of platelet accumulation (green) and coagulation (violet), and (C) moment-of-onset are shown as functions of platelet concentrations in reconstituted blood. Dots are mean values, whiskers are standard deviations. Please click here to view a larger version of this figure.
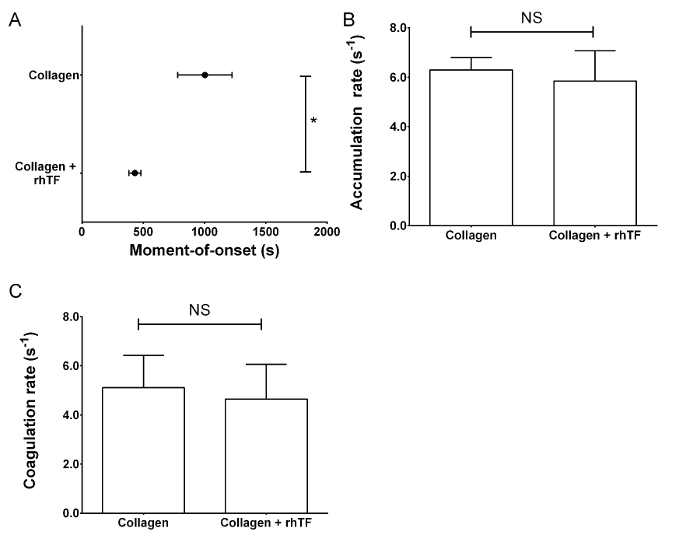
Figure 4: Activation of both TF and contact pathway coagulation significantly shortens the moment-of-onset of coagulation. Reconstituted blood, in the presence of Ca2+, was perfused through chambers coated with collagen alone or with collagen and rhTF to study the contact pathway alone or a combination of the contact and TF pathways. (A) The moment-of-onset of coagulation is significantly shortened in TF-containing flow chambers. (B) The rate of accumulation of platelets during coagulation and (C) the rate of coagulation are not significantly different between collagen only or collagen- and rhTF-coated flow chambers. Bars and dots are mean values; whiskers are standard deviations. Statistical analysis was by paired t-test. (NS = not significant, *P <0.05, n ≥3). Please click here to view a larger version of this figure.

Supplementary Video 1: In the absence of TF, coagulation in collagen-coated perfusion flow chambers depends on FXIIa. The role of the contact pathway is determined in collagen-coated perfusion flow chambers. (A) Overlaid fluorescence image sequence of platelet (green) and fibrin(ogen) (violet) deposition in the absence of CTI. (B) Same sequence as panel A but with the green channel switched off. (C) Overlaid fluorescence image sequence of platelet (green) and fibrin(ogen) (violet) deposition in the presence of CTI. (D) Same sequence as panel C but with the green channel switched off. Please click here to download this video.
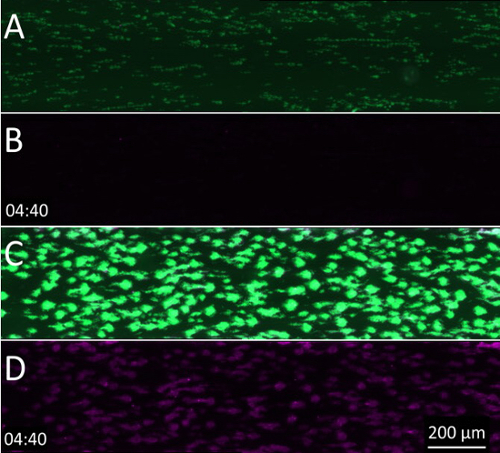
Supplementary Video 2: Activation of both TF and the contact pathway significantly shortens the coagulation moment-of-onset. To study the role of TF, flow chambers coated with collagen alone or with collagen and rhTF were used. (A) Overlaid fluorescence image sequence of platelet (green) and fibrin(ogen) (violet) deposition in flow chambers coated only with collagen. (B) Same sequence as panel A but with the green channel switched off. (C) Overlaid fluorescence image sequence of platelet (green) and fibrin(ogen) (violet) deposition in flow chambers coated with both collagen and rhTF. (D) Same sequence as panel C but with the green channel switched off. Please click here to download this video.
