To obtain the results shown in this method, the open reading frame of green fluorescent protein (GFP) is fused to a promoter/wound-induced enhancer sequence from Dopa Decarboxylase (Ddc) and DsRed is fused to a wound enhancer sequence from tyrosine hydroxylase (ple)4,6. In a live Drosophila embryo, maturation of the fluorescent wound reporter protein is optimal 4-6 hr after wounding, which allows time for the accumulation of sufficient protein to visualize, as well as time for the fluorescent protein to oxidize to the fluorescent state15. To observe the reporter localization, a compound fluorescent microscope is sufficient. To observe a higher magnification of the reporter localization, a confocal microscope is optimal to generate a maximum projection of multiple optical sections (Figure 1). Confocal in vivo imaging of Drosophila embryos is complicated by the rapid undulations of the embryo. . A full-view of embryo can be obtained with 20X objective and a close-up view of wound site can be obtained with 40X objective. A top-view of epidermal wound reporter localization can be imaged with 10 successive 1µm optical sections. A lateral-view of epidermal wound reporter localization can be imaged with 40 successive 1 µm optical sections.
Using standard genetic crossing methods, it is possible to combine wound reporters with genetic mutations. One example presented in this paper is Flotillin-2 (Flo-2), because loss-of-function (null allele generated with P-element insertion) and gain-of-function (overexpression generated with ubiquitous expression in all cells) mutants of Flo-2 demonstrate the Flo-2 gene product is necessary and sufficient to inhibit the localization of the Ddc-GFP wound reporters9 (Figures 2A and 2B). Using the microinjection protocol, it is possible to test whether introduction of chemical solutions superactivates or inhibits the localization of wound reporters. Chemically-wounded embryos were simultaneously wounded and injected with a 1:4 ratio of 1% toluidine blue dye and solubilized compounds. Toluidine blue dye allowed for visual confirmation of solubilized compounds being injected into the body cavity. Control embryos were wounded with a broken needle containing 1:4 ratio of 1% toluidine blue dye and solute without chemical. Two examples presented in this paper are hydrogen peroxide (H2O2) and methyl-β-cyclodextrin (MβCD), because both chemical solutions are sufficient to globally activate the localization of the Ddc-GFP wound reporters9 (Figures 2C and 2D). Hydrogen peroxide (H2O2) was diluted in H2O to 0.6 M. Methyl-β-cyclodextrin (MβCD) was solubilized in 1 mM NaOH to 3 mM. Using the embryo fixation protocol, it is possible to detect the transcription activation of wound response genes in a localized domain, surrounding a wound site. One example presented in this paper is the in situ hybridization of RNA probes to detect Ddc and ple transcripts4,6,9 (Figures 3A and 3B).
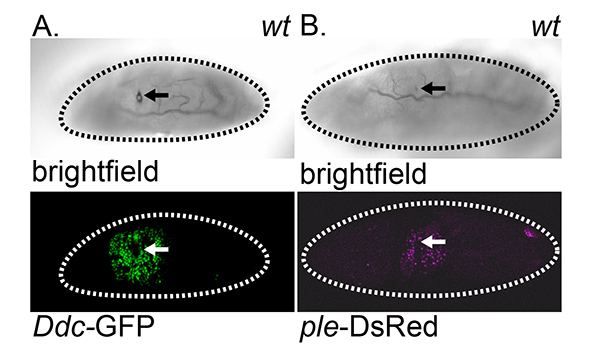
Figure 1. Ddc-GFP and ple-DsRed wound reporter localization. Brightfield and fluorescent images in wounded wild type embryos. (A) Top-view of Ddc-GFP wound reporter. (B) Top-view of ple-DsRed wound reporter. All images were collected on a Leica SP2 confocal microscope, with 20X objective. Arrows mark site of wound. Dashed lines in the data panels mark the outlines of embryos. Click here to view larger image.

Figure 2. Localization of the Ddc-GFP wound reporter in genetic mutant backgrounds and chemical microinjected embryos. Brightfield and fluorescent images of wounded Flotillin-2 (Flo-2) mutant embryos. (A) Loss-of-function Flo-2 {KG00210} mutants, expansion of reporter activity throughout all epidermal cells. (B) Gain-of-function Flo-2 (armadillo-GAL4, UAS-Flo2) mutants, inhibition of reporter activity throughout all epidermal cells. (C) Hydrogen peroxide (H2O2) solution microinjection, expansion of reporter activity throughout all epidermal cells. (D) Methyl-β-cyclodextrin (MβCD) solution microinjection, expansion of reporter activity throughout all epidermal cells. All images were collected on a Leica SP2 confocal microscope, with 20X objective. Arrows mark site of wound. Dashed lines in the data panels mark the outlines of embryos. Click here to view larger image.
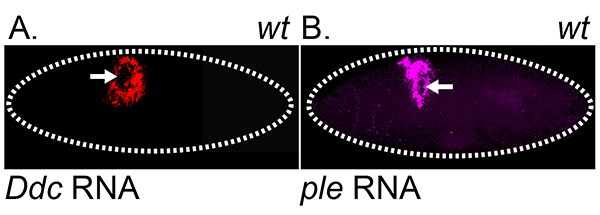
Figure 3. Detection of wound response gene RNA transcripts. Fluorescent images of in situ hybridization and RNA detection in wounded wild type embryos. Ddc (A) and ple (B) RNA transcripts accumulate around the wound site. All images were collected on a Leica SP2 confocal microscope, with 20X objective. Arrows mark site of wound. Dashed lines in the data panels mark the outlines of embryos. Click here to view larger image.
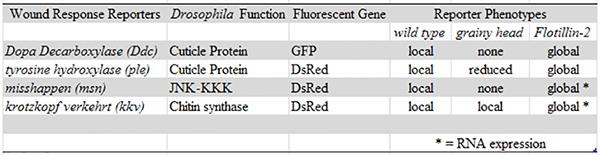
Table 1. Summary of epidermal wound response reporters. Insertions of the four wound response reporters (Ddc, ple, msn, kkv) are available on both the second and third chromosomes. All of the wound response reporters activate the fluorescence gene (GFP or DsRed) in a limited number of epidermal cells surrounding the site of puncture injury in the wild type background (e.g. "local" phenotype). Ddc and msn wound response reporters require Grh for activation of the fluorescence gene after puncture injury (e.g. "none" phenotype). All of the wound reporters require Flo-2 for localization of the fluorescence gene after puncture injury (e.g. "global" phenotype). Click here to view larger image.
