To observe the efficiency of the technique detecting variations in chlorophyll cellular concentration and evaluate the effect of growth factors in C. sorokiniana, a Taguchi L4 experimental design was established, evaluating CO2 volume addition, light color, and light intensity. Each factor was assessed at low and high levels, as shown in Table 1, under the conditions defined by the experimental design in Table 2.
Once the experimental process was concluded, the results showed that the combination of low addition of CO2, purple light, and high light intensity (C1, P, I2) corresponding to treatment 2, increased microbial growth to 7.83 x 105 cells mL-1. On the other hand, chlorophyll content was higher in treatment 4 under conditions C2, P, I2, reaching a concentration of 5.92 x 10-7 µg cel-1 at 132 h (Figure 1) coinciding with purple light and high light intensity as the optimal levels of the factors in both experiences.
At the same time, a similar behavior was observed across the four evaluated lines. However, a clear favorability was observed in the tests conducted under purple light. This pattern suggests that purple light may positively impact the outcomes, potentially promoting optimal conditions for cellular growth or activity. These findings underscore the importance of carefully considering lighting conditions in biological experiments.
From the statistical analysis, the main effects plot was obtained. In Figure 2, it can be observed how each level of the evaluated factors influences chlorophyll production in C. sorokiniana. The best conditions for chlorophyll production were high addition of CO2, purple light, and high light intensity (C2, P, I2), where color and carbon dioxide concentration show very close significance values, meanwhile light intensity shows a lower significant effect on the evaluated response variables.
In addition, a contrast diagram was generated (Figure 3), showing the behavior of cellular chlorophyll concentration according to the effect of CO2 volume addition and the light color to which they were subjected.
It was observed that purple light (a mixture of blue and red light) and a high light intensity significantly promote microbial growth and chlorophyll production. Meanwhile, a high addition of CO2 influences both parameters positively and negatively, respectively, as it increases chlorophyll cellular concentration to a maximum during the experimentation but hinders microbial growth. However, it is noticeable that under purple light, a CO2 volume addition of 20 s pulses of CO2 every 12 h at a flow rate of 8.5 ft³ h-1 allows a favorable response for both parameters.
To validate the method and results obtained from C. sorokiniana, a confirmatory flask-level test was conducted on H. pluvialis under red light due to its natural characteristic of degrading chlorophyll to synthesize astaxanthin under stress conditions, which were induced at 168 h by increasing the salinity of the medium. A concentration graph was generated (Figure 4) where the maximum chlorophyll concentration was obtained at 0 h with 1.41 x 10-7 µg cel-1. In this case, due to the natural microalgae behavior, a decrease in chlorophyll concentration was observed, suggesting potential physiological changes in the biological model under investigation. This is further illustrated in Figure 5, which shows a micrograph of C. sorokiniana (Figure 5A) and two micrographs of H. pluvialis: one in the green phase (Figure 5B) and one in the phase of change and astaxanthin production (Figure 5C). Interestingly, despite this decline, no interference occurred from the presence of carotenoid compounds, which are, in turn, fat-soluble pigments present in them; this is strong evidence of the selectivity and reproducibility of the technique in microalgae with diverse capacities and behaviors.
The graph depicts a decrease in total chlorophyll concentration throughout the experiment. Notably, there is an increase in chlorophyll a compared to chlorophyll b at 50 h, where the concentration of chlorophyll b decreases significantly. The most substantial decline in chlorophyll concentration occurs at the end of the exponential growth phase. This decrease is attributed to the production of carotenoids in H. pluvialis.
Similarly, the analysis of chlorophyll a and b composition was conducted for the treatment with the highest production of C. sorokiniana (C1, P, I2) in Figure 6. Contrary to H. pluvialis, the production of chlorophylls displayed a consistent increase throughout the entire experimentation period, which was solely analyzed during the exponential phase. It is observed that similar to H. pluvialis, chlorophyll a is initially present in lower proportions than chlorophyll b at the beginning of the experiment; however, these proportions undergo a notable shift at the 50 h mark, with chlorophyll a showing a higher production rate than chlorophyll b thereafter.
The analytical technique employed in this study has proven to be efficient in detecting changes within a biological model and across different biological models. These findings underscore the sensitivity and precision of the analytical method, allowing for a nuanced understanding of chlorophyll dynamics within and between biological systems. Such insights are invaluable for unraveling the intricate mechanisms governing pigment synthesis and metabolism, paving the way for further advancements in biotechnological and environmental research.
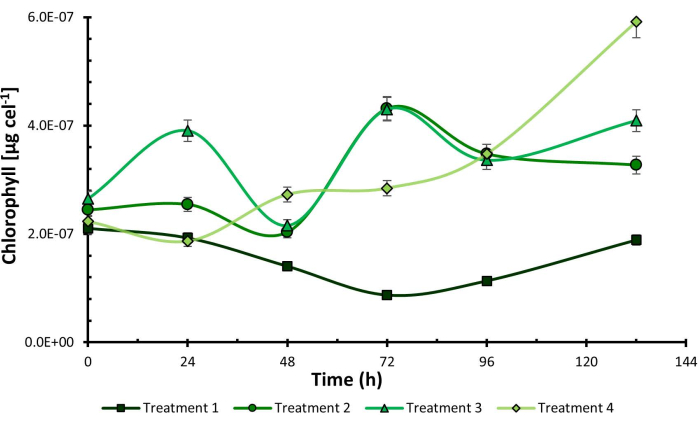
Figure 1: Chlorophyll cellular content under experimental design treatments. Chlorophyll cellular content measured in Chlorella sorokiniana across different experimental treatments. Data presented as standard deviation for n = 3. Please click here to view a larger version of this figure.
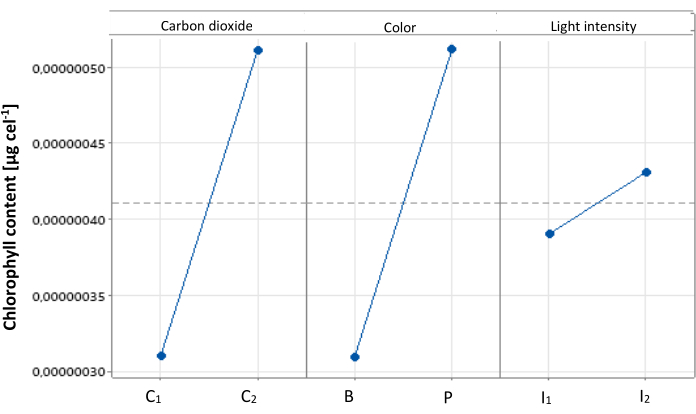
Figure 2: Main effects plot for cellular chlorophyll content. Main effects plot illustrating the impact of different treatments on cellular chlorophyll content in Chlorella sorokiniana. Data presented as standard deviation for n = 3. Please click here to view a larger version of this figure.
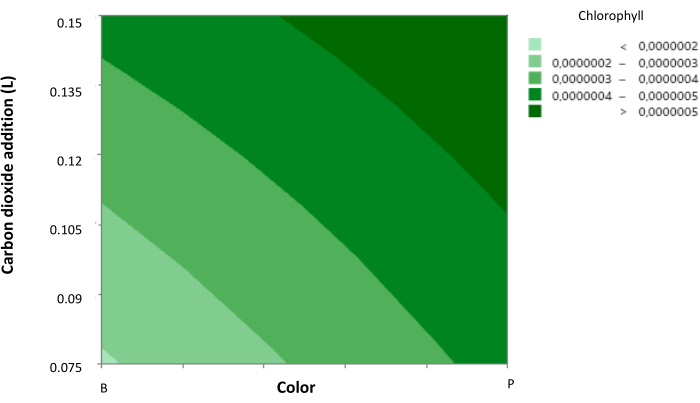
Figure 3: Contour plot of cellular chlorophyll concentration. Contour plot showing the relationship between cellular chlorophyll concentration, CO2 concentration, and light color. Light color is represented with axis 1 as blue and axis 2 as purple, with intermediate levels indicating the addition of red light. Data presented as standard deviation for n = 3. Please click here to view a larger version of this figure.
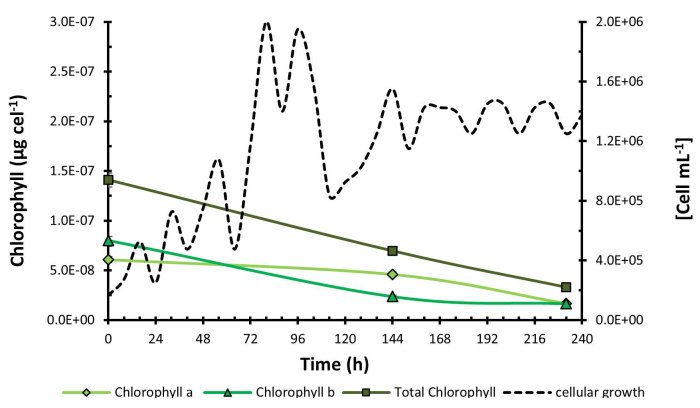
Figure 4: Chlorophyll concentration during H. pluvialis growth kinetics. Chlorophyll concentration measured over the growth kinetics of Haematococcus pluvialis. Data presented as standard deviation for n = 3. Please click here to view a larger version of this figure.
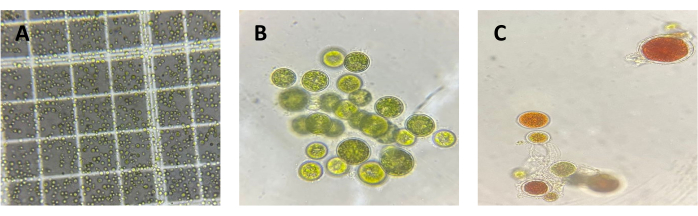
Figure 5: Micrographs of C. sorokiniana and H. pluvialis. Micrograph images of Chlorella sorokiniana (A) and Haematococcus pluvialis, illustrating one specimen in the green phase (B) and another in the early red phase (C). Please click here to view a larger version of this figure.
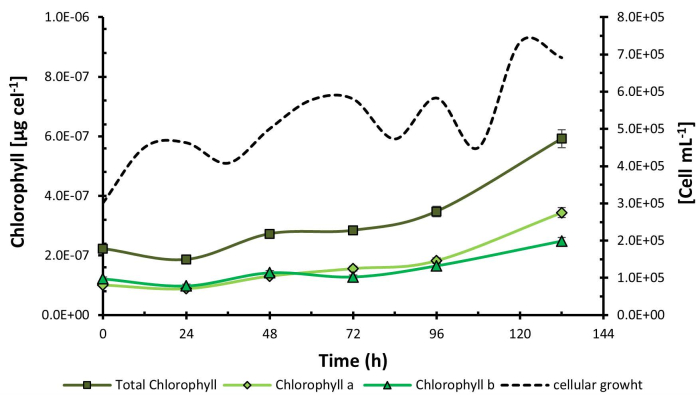
Figure 6: Chlorophyll concentration during C. sorokiniana growth kinetics. Chlorophyll concentration measured over the growth kinetics of Chlorella sorokiniana. Data presented as standard deviation for n = 3. Please click here to view a larger version of this figure.
| Factor | Low level | High level |
| CO2 addition (L) | C1 | C2 |
| Light color | Blue (B) | Purple (P) |
| Light intensity (Lux) | I1 | I2 |
Table 1: Kinetics factors and levels. Overview of kinetics factors and their respective levels: C1 = 0.078 L; C2 = 0.15 L; B = Blue light; P = Purple light; I1 = 550 lux; I2 = 1200 lux.
| Treatment | CO2 addition | Light color | Light intensity (Lux) |
| (L) | |||
| 1 | C1 | B | I1 |
| 2 | C1 | P | I2 |
| 3 | C2 | B | I1 |
| 4 | C2 | P | I2 |
Table 2: Experimental design and treatments. Summary of the experimental design and treatments applied: C1 = 0.078 L; C2 = 0.15 L; B = Blue light; P = Purple light; I1 = 550 lux; I2 = 1200 lux.
Supplementary File 1: Cellular quantification data. Please click here to download this File.
