A previously published dataset of leaf physiognomy measurements from the early Eocene McAbee fossil site in south-central British Columbia was used to provide an example of representative results using both the digital leaf physiognomy (DiLP) and leaf mass per area (MA) reconstruction methods (Lowe et al.38; data provided in Supplementary File 2). The site provides an opportunity to reconstruct paleoclimate and paleoecology during the warmest interval of the Cenozoic (the Early Eocene Climatic Optimum) in an upland and volcanic landscape38,45,46,47. Fossil assemblages were sampled from two separate horizons in a lacustrine sequence, named H1 (28 cm thick) and H2 (27 cm thick), pooled over a narrow range of stratigraphy using a census technique, whereby all specimens able to be assigned a morphotype were collected or counted38,48.
The McAbee leaf physiognomic data passed the error checks flagged by dilp_errors(), and seven outliers flagged by dilp_outliers() were double-checked to ensure the values represent true variation in data and not a methodological mistake. The data was subsequently run through the dilp() function to produce paleoclimate and the lma() function for leaf mass per area reconstructions.
MA reconstructions and the lower and upper bounds of their 95% prediction intervals are reported in Table 4 at both the species- and site level, using equations presented in Royer et al.5 and Butrim et al.10. Reconstructed values are within the range of MA typical for modern terrestrial species (30-330 g/m2)49. Using thresholds discussed in Royer et al.5, most species have a reconstructed MA that aligns with leaf life spans of <1 year (≤87 g/m2), some ~1 year (88-128 g/m2), while none are typical of >1 year (≥129 g/m2). Reconstructions of site MA mean, and variance at McAbee reflect the prevalence and diversity of leaf economic strategies at a site10,50. There are no prominent differences between site mean and variance between H1 and H2, and thus, there is no evidence that the composition and diversity of leaf economic strategies varied between the two points in time. Additionally, the site-mean reconstructions made using the equations of Royer et al.5 and Butrim et al.10 were very similar.
Reconstructions of mean annual temperature (MAT) and mean annual precipitation (MAP) using multiple linear regression (DiLP) and single linear regression (leaf margin and leaf area analyses) equations presented in Peppe et al.6 are shown in Table 5. Paleoclimate estimates are most reliably inferred if the leaf physiognomy of the fossil leaf assemblages occurs within the physiognomic space of the calibration dataset. This is assessed through the canonical correspondence analysis (CCA) analysis step carried out by the function dilp_cca(). Both McAbee H1 and H2 fall within the range of leaf physiognomy observed in the calibration dataset (Figure 7A). If sites had reconstructed values that fell outside calibration space, paleoclimate reconstructions should be interpreted cautiously (e.g., through comparison to independent lines of evidence; see Peppe et al.6 for further discussion). Reconstructed MAT and MAP for both H1 and H2 are consistent with a temperate seasonal biome (Figure 7B,C), which agrees well with independent lines of evidence, including nearest living relative based inferences of both the fossil floral and insect communities at McAbee45.

Figure 1: Leaf physiognomy and architectural terminology throughout this article. (A) A pinnately veined, unlobed, and entire-margined leaf, (B) a palmately veined, unlobed, and toothed leaf, (C) a pinnately veined, lobed, and entire-margined leaf, (D) a palmately veined, lobed, and toothed leaf. Please click here to view a larger version of this figure.
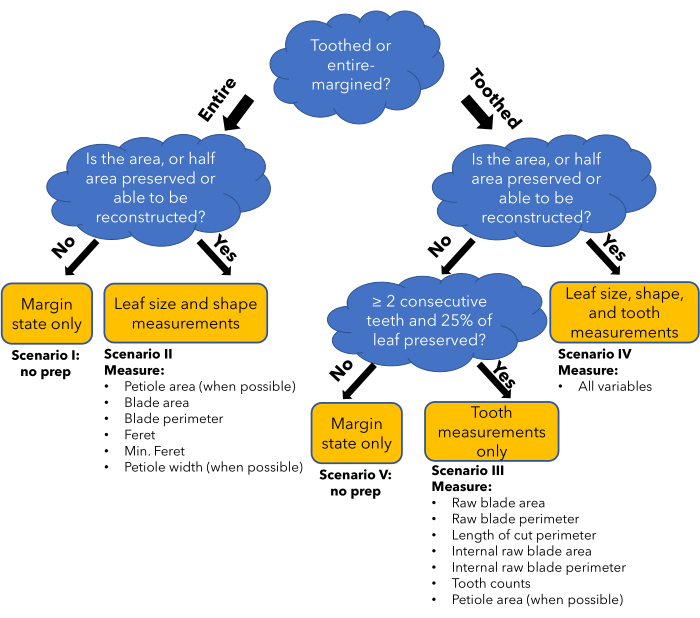
Figure 2: Flow chart of the method. A flowchart demonstrating how different leaf preservation conditions and leaf types determine what general type of leaf traits can be measured reliably (yellow box). This determines which preparation scenario will be followed in the protocol, and in which columns data will be entered in the data entry spreadsheet (bullet points). Please click here to view a larger version of this figure.

Figure 3: Different preparation scenarios. Different preparation scenarios demonstrating examples of completed digitally prepared images ready for the measurement phase. (A) Scenario 1, entire margined leaf whose area, or half area, cannot be reconstructed, (B) Scenario 5, toothed leaf whose area, or half area, cannot be reconstructed and does not have ≥2 consecutive teeth and/or ≥25% of the leaf preserved, (C) Scenario 2, entire margined leaf whose area, or half area, is preserved or can be reconstructed, (D) Scenario 3, a toothed leaf whose area, or half area, cannot be reconstructed but has ≥2 consecutive teeth and ≥25% of the leaf preserved, (E) Scenario 4, a toothed leaf whose area, or half area, is preserved or can be reconstructed. Please click here to view a larger version of this figure.

Figure 4: Illustration of damage removal. Illustrating how to cut out damaged margin, and the leaf area adjacent to that damaged margin. Dashed red lines demonstrate how selections are made with the lasso tool. Note that the bounds of damage were intentionally started at the sinuses of primary teeth (see Supplementary Figure 2 for help differentiating primary from subsidiary teeth). (A) A pinnately veined leaf where the selection is extended to the mid-vein. (B) A palmately veined leaf where the selection is extended to the nearest primary vein. Please click here to view a larger version of this figure.
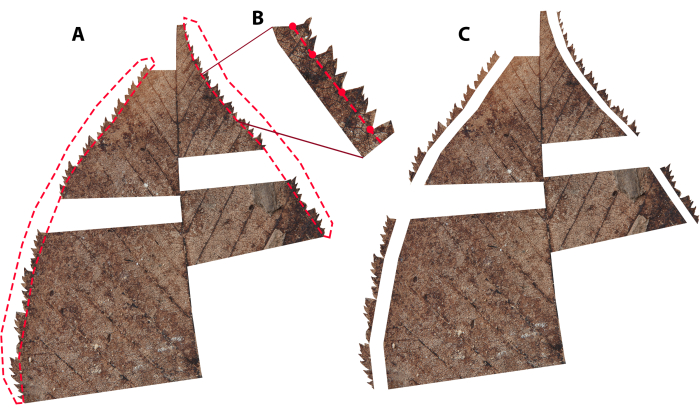
Figure 5: Illustrating an example of how to cut out teeth. (A) Dashed red lines demonstrate how selections are made with the lasso tool. Note that in this case, the teeth are compound, so selections were made between primary sinuses only (see Supplementary Figure 2 for help differentiating primary from subsidiary teeth), (B) a zoomed-in perspective of how teeth selection was made, with red dots representing where the mouse was clicked during selection, (C) the copy of the leaf when the teeth are removed. Please click here to view a larger version of this figure.
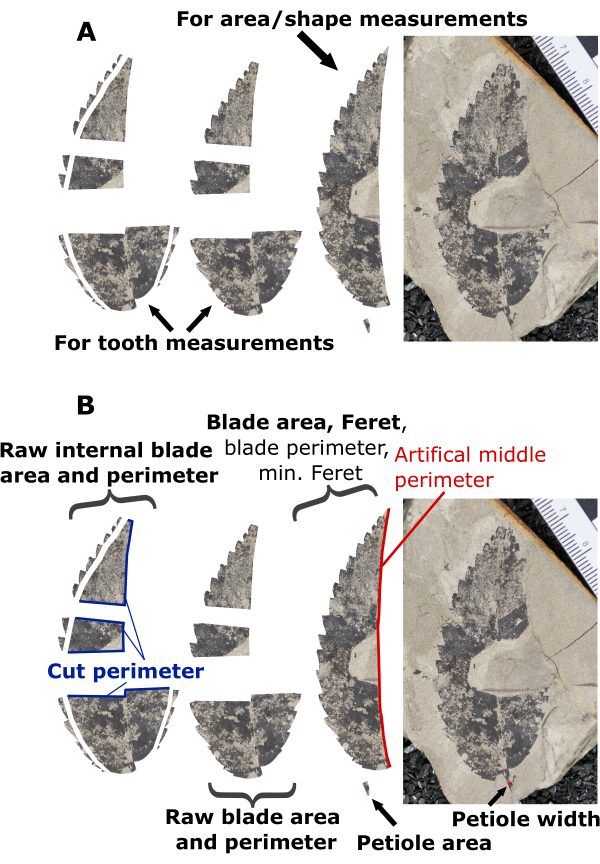
Figure 6: Illustration of preparation scenario 4. Illustration of preparation decisions and measuring steps for an example leaf prepared in scenario 4. (A) A preparation scenario where it was decided that a half leaf provided the most reliable leaf shape and area measurements, and preserved margins on both medial halves were included for tooth measurements. (B) An example demonstrating which variables are measured on various components of the prepared leaf. Bolded text highlights measurements needed for DiLP and MA analyses, while non-bolded text (blade perimeter, minimum Feret, and artificial middle perimeter) highlights measurements that are not required but useful for additional physiognomic characterizations (e.g., shape factor and compactness; Table 1). Please click here to view a larger version of this figure.
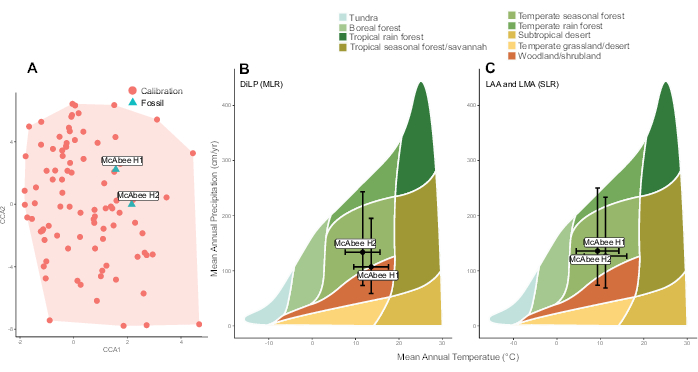
Figure 7: Representative results. Results from two fossils horizons (H1 and H2) sampled at the early Eocene McAbee Fossil Beds from Lowe et al.38. (A) Canonical correspondence analysis showing the representation of multivariate leaf physiognomy in the calibration dataset. Calibration data is from Peppe et al.6. The leaf physiognomy of the two McAbee horizons are overlain and occur within the calibration space. (B and C) Temperature and precipitation estimates, and their associated uncertainty (standard errors of the models), using equations presented in Peppe et al.6 of the two McAbee horizons overlain on a Whittaker Biome diagram. (B) Estimates reconstructed using the Digital Leaf Physiognomy (DiLP) multiple linear regressions models (MLR), (C) Estimates reconstructed using the leaf area analysis (LAA) and leaf margin analysis (LMA) single linear regressions (SLR) equations of the two McAbee horizons overlain on a Whittaker Biome diagram. Please click here to view a larger version of this figure.
Table 1: Leaf physiognomic variables. Variables that are measured and/or calculated and applied in predictive models using this protocol to reconstruct leaf dry mass per area (MA), mean annual temperature (MAT), and mean annual precipitation (MAP). MAT and MAP are reconstructed with equations presented in Peppe et al.6 using a multivariate approach for Digital Leaf Physiognomy (DiLP) and univariate approaches for leaf margin analysis (LMA) and leaf area analysis (LAA). Variables listed as Other are not used in MA, DiLP, LMA, and LAA analyses but are still measured and calculated using this protocol because they are easy to implement and provide useful characterizations of leaf physiognomy. Please click here to download this Table.
Table 2: Additional considerations and explanations for preparation steps. Please click here to download this Table.
Table 3: Additional considerations and explanations for measuring steps. Please click here to download this Table.
Table 4: Reconstructions of leaf dry mass per area (MA) and associated upper and lower bounds of the 95% prediction intervals for McAbee Fossil Beds from Lowe et al.38. Reconstructions are made for morphotype mean5, site mean5,10, and site variance10. Please click here to download this Table.
Table 5: Reconstructions of mean annual temperate (MAT) and mean annual precipitation (MAP) for Horizon 1 (H1) and 2 (H2) at the early Eocene McAbee Fossil Beds using the multiple linear regressions (MLR) of Digital Leaf Physiognomy (DiLP) and the single linear regressions (SLR) of leaf margin analysis (LMA) and leaf area analysis (LAA) presented in Peppe et al.6. Please click here to download this Table.
Supplementary Figure 1: Quercus rubra leaf from Harvard Forest illustrating the lobe vs. tooth rule. Line segments p and d are defined in text. Scale bars = 1 cm. Please click here to download this File.
Supplementary Figure 2: Betula lutea leaf from Harvard Forest illustrating the rules for differentiating subsidiary teeth from primary teeth. The isolated leaf segment has been magnified 2X. The blue line connects sinuses with the greatest degree of incision (i.e., primary sinuses), and teeth associated with these sinuses are considered primary (blue arrows). Red dots mark teeth that can be differentiated as a subsidiary because their apical sinuses are incised to a lesser degree. Teeth denoted by the red arrows have a similar degree of incision compared to the primary teeth but can be identified as subsidiary by a relatively thinner gauged principal vein compared to the primary teeth. Scale bars = 1 cm. Please click here to download this File.
Supplementary Figure 3: Illustration of tooth selection, the pinnate lobe rule, and the lobe priority rule. (A) Tooth selection for a Hamamelis virginiana leaf from Huyck Preserve. The darkened areas correspond to leaf tissue that is included in total tooth selection because subsidiary teeth are differentiated from primary teeth. (B) Quercus alba leaf from IES illustrates the lobe priority rule. The darkened areas are measured as lobes, and the undarkened are measured as teeth, but all projections are considered lobes via the lobe priority rule. Scale bars = 1 cm. Please click here to download this File.
Supplementary Figure 4: Acer saccharum leaf from Allegheny National Forest illustrating the extension and solitary tooth rules. Dashed lines depict tooth selections. The solid line depicts the axis of symmetry for the associated tooth. The black area is a weight used to flatten the leaves for photography. Scale bars = 1 cm. Please click here to download this File.
Supplementary Figure 5: Illustrating the ideal way to cut a petiole out that is positioned on top of a cordate base. Please click here to download this File.
Supplementary File 1: Data entry template for all measured digital leaf physiognomy variables. This file should not be modified, as it will be used as the input file for the R package. Please click here to download this File.
Supplementary File 2: Example data from McAbee fossil beds from Lowe et al.38. This data was used to generate Figure 7 and for the discussion of representative results. Please click here to download this File.
Supplementary File 3: Rules document for fossil digital leaf physiognomy. Please click here to download this File.
