The use of a chromatographic column-based protocol allows the processing of significant volumes of HEK293 conditioned media. In Figure 1, the results of the purification of BDNFAvi from 500 mL of conditioned media are shown. Consecutive elutions of BDNFAvi from the Ni-NTA agarose beads yield decreasing concentrations of BDNFAvi (Figure 1A). After four consecutive elutions (each lasting 15 min), the majority of the BDNF captured by the beads is recovered. The concentrations of the eluates range from 6 to 28 ng/μL, and the total yield amounted to approximately 60 μg of BDNFAvi (Table 1). The produced BDNFAvi was then efficiently biotinylated by an in vitro reaction mediated by BirA-GST, as demonstrated by the lack of non-biotinylated BDNFAvi in the supernatant (Figure 1B). Please note that the biotinylation presented in Figure 1B corresponds to an aliquot of the total BDNF produced, but the reaction can be scaled up for bigger volumes.
Then, the biological activity of mbtBDNF was evaluated using 2 different experimental approaches. First, cortical neurons seeded in 60 mm plates (2 million neurons, DIV7) were stimulated with 50 ng/mL of mbtBDNF for 30 min, and then proteins were prepared for western blot analysis. The biological activity of the mbtBDNF was quantified by detecting pTrkB (Y515) and pERK (T202/Y204). Binding of BDNF to TrkB triggers the activation of the receptor through an autophosphorylation reaction in its intracellular domain, and ERK is a known target of the BDNF signaling pathway22. The bands for both phosphorylated proteins had a similar intensity in neurons treated with commercial BDNF and mbtBDNF, and both showed a stronger signal than control condition (Figure 2A). Then, the biological activity of mbtBDNF coupled to streptavidin-QD was evaluated to demonstrate that they can be used in live imaging experiments. Cortical neurons were seeded in 10 mm covers (40,000 cells per cover, DIV7) and treated with a final concentration of 200 pM or 2 nM BDNF-QD for 30 min before fixing and staining for pCREB. CREB is a transcription factor which is targeted by activated ERK1/2 in cortical neurons22,23. Stimulating neurons with increasing concentrations of BDNF-QD resulted in a dose-dependent increase of phosphorylation of CREB and presence of QD particles surrounding the nucleus (Figure 2B), indicating that the BDNF-QD particles were endocytosed and triggered the activation of signaling pathways associated with BDNF-mediated TrkB activation. A twofold increase in pCREB signal was detected when stimulating neurons with a low concentration of BDNF-QD (200 pM), whereas stimulating with 2 nM resulted in a 3.5-fold increase in the pCREB signal (Figure 2C). These results demonstrate that the biotinylated BDNFAvi is biologically active, and that it does not lose its activity when coupled to streptavidin-QD, making it suitable for immunofluorescence and live cell imaging.
Finally, the imaging potential of BDNF-QD was evaluated in compartmentalized cultures using microfluidic chambers. Cortical neurons were seeded in microfluidic chambers (15 mm covers, 50,000 neurons per microfluidic chamber, DIV7) to separate the axonal and somatodendritic compartments and were stimulated with 2 nM BDNF-QD for 3.5 h. Live cell microscopy was performed, and the resulting kymographs were used to quantify the speed of BDNF-QD containing organelles (Figure 3A). An average moving speed of 0.91 μm/s was detected (Figure 3B), which is in line with previous analyses of cytoplasmic dynein-mediated transport7,16. Microfluidic chambers treated with 2 nM streptavidin-QD did not show moving QDs in the microgrooves, as shown by the kymograph (Figure 3A). Cells grown under the same conditions were stimulated with 500 pM or 2 nM BDNF-QD for 210 min, and then fixed and labelled with a nuclear staining. As shown in Figure 3C, neurons show a dose-dependent accumulation of BDNF-QD in all the analyzed sub-compartments, including the proximal and distal portions of the microgroove and the somatodendritic compartment. In contrast, control neurons showed almost no QD signal throughout the chamber. Therefore, the BDNF-QD can be detected in live and fixed cells in microfluidic chambers.
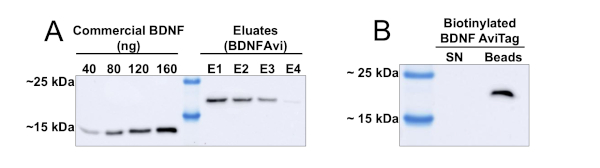
Figure 1: Production and mono-biotinylation of BDNFAvi in HEK293 cells. HEK293 cells were transfected using the PEI reagent and a BDNFAvi encoding plasmid and the conditioned media was collected after 48 h. BDNFAvi contains a 6x Histidine tag allowing purification using nickel-nitrilotriacetic acid (Ni-NTA) chromatography. Commercially available recombinant human BDNF has an expected molecular weight of ~13 kDa, whereas BDNFAvi displays a molecular weight of ~18 kDa. BDNFAvi bound to the resin was fully eluted with four consecutive elution steps. (A) Western blot using anti-BDNF antibodies to detect in house prepared recombinant BDNF and commercial BDNF. Aliquots containing known amounts of commercially available human BDNF and 5 μL of each eluate were loaded into an SDS-PAGE gel for detection of BDNFAvi using an antibody against BDNF. Table 1 indicates the concentrations of BDNFAvi present in each eluate. The amount and concentration of BDNF in each eluate was obtained by densitometric analysis and interpolation from the concentration curve of commercially available BDNF. (B) Verification of BDNFAvi biotinylation. Eighty nanograms of biotinylated BDNFAvi (mbtBDNF) were incubated with 30 μL of streptavidin coupled to magnetic beads (20% slurry) for 1 hr at 4 °C. Then, magnetic beads were isolated using a magnetic separator. The streptavidin beads were heated with loading buffer to elute the biotinylated BDNFAvi (beads lane). The supernatant (SN lane) was also treated with loading buffer, heated and loaded in the gel (SN lane). Please click here to view a larger version of this figure.
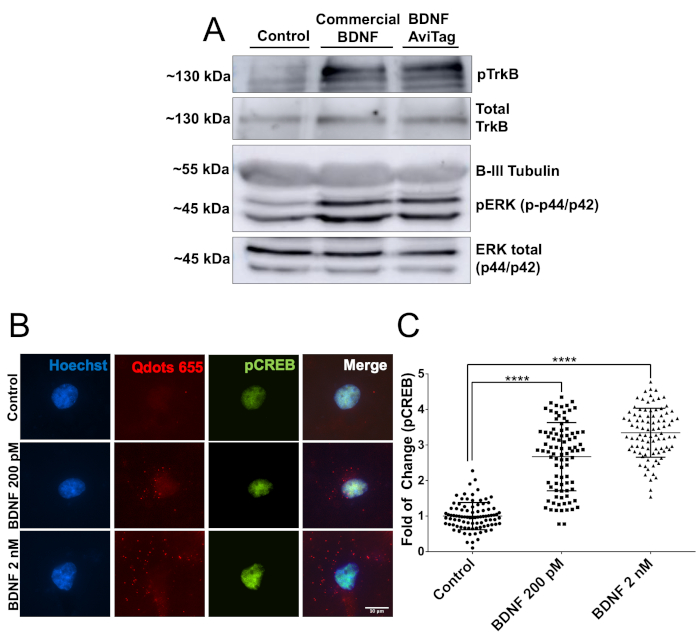
Figure 2: Verification of mbtBDNF biological activity. (A) DIV7 cortical neurons were serum starved for 1 h, and then stimulated with 50 ng/mL of commercially-available BDNF or mbtBDNF for 30 min. Proteins were extracted and loaded in an SDS-PAGE gel for analysis of TrkB and ERK1/2 phosphorylation using phospho-specific antibodies and compared to the total levels of the protein using antibodies against total TrkB and ERK1/2. (B) DIV7 cortical neurons were serum starved for 1 h, and then stimulated with a final concentration of 200 pM or 2 nM of mbtBDNF coupled to streptavidin-QD (BDNF-QD) for 30 min. Then, cells were fixed and pCREB was labelled for fluorescence microscopy analysis. (C) Quantification of nuclear pCREB fluorescence intensity. The results correspond to 90 neurons pooled together from 3 independent experiments, shown as mean ± SEM. The statistical analysis corresponds to a one-way ANOVA with Tukey’s multiple comparisons test (****p < 0.0001). Please click here to view a larger version of this figure.
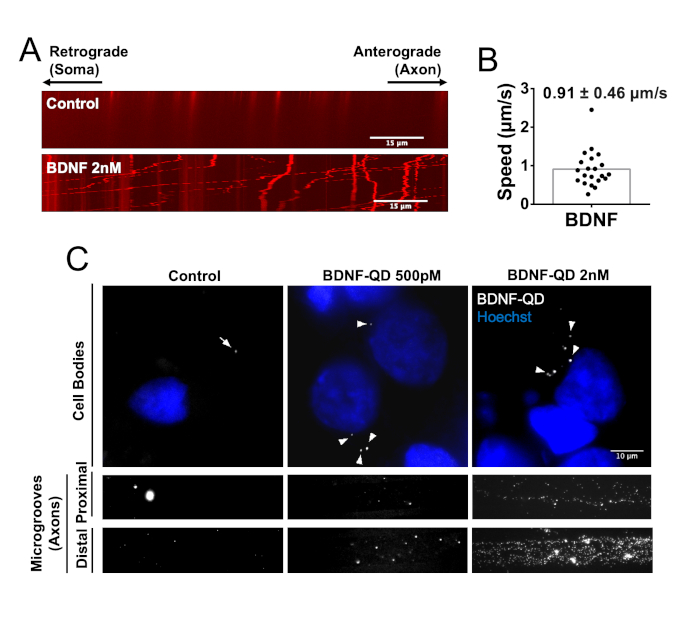
Figure 3: Visualization of BDNF-QD in live and fixed cells. (A) DIV7 cortical neurons grown in microfluidic chambers were stimulated in the axonal compartment with a final concentration of 2 nM BDNF-QD for 3.5 hrs, and then the proximal portion of the microgrooves was imaged using a live cell microscopy setting. Representative kymographs for control condition (treated with streptavidin-QD) and upon treatment with BDNF-QD are shown. (B) Quantification of the speed of moving BDNF-QD. Mobile puncta were defined as those that moved more than 10 μm in the 120 s of recording. (C) DIV7 cortical neurons grown in microfluidic chambers were stimulated in the axonal compartment with a final concentration of BDNF-QD of 500 pM or 2 nM for 3.5 hrs, and then fixed and labelled with Hoechst to visualize the nuclei. Representative images of the somatodendritic compartment and the distal and proximal portions of the microgrooves are shown. Please click here to view a larger version of this figure.
| Eluate | BDNF (ng) in 5μl | [ng/μl] | Total BDNF [μg] |
| E1 | 142.2 | 28.4 | 28.4 |
| E2 | 101.2 | 20.2 | 20.2 |
| E3 | 65.6 | 13.1 | 13.1 |
| E4 | 30.4 | 6.1 | 6.1 |
| 67.8 |
Table 1: Quantification of BDNFAvi purification yield (related to Fig. 1A). HEK293 cells were transfected with a plasmid driving BDNFAvi expression, and the protein was purified by Ni-NTA affinity chromatography. Protein concentration and final yield was calculated by densitometric analysis and interpolation in the known concentration curve of commercially available recombinant human BDNF.
Supplemental File 1: Culture media and buffer components Please click here to download this file.
