All animal work utilizes protocols reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) at Geisel School of Medicine at Dartmouth.
1. Collection of fluids
NOTE: Both serum and CSF are required. Two protocols for each fluid collection are needed for survival and necropsy.
- Serum and CSF collection using survival procedures
NOTE: For survival fluid collection, serum collection should precede CSF collection as it is a less invasive procedure. CSF must be obtained within one week of serum draw.- Retro-orbital bleeding procedure for serum collection.
NOTE: This procedure is for survival bleeding of mice14. The procedure described applies to any age, gender, and strain of mice. Since IACUC rules dictate that a maximum blood volume of 1% of body weight can be removed as a single blood draw, it is recommended the procedure is performed only on mice weighing more than 15 g.- Move the cages containing mice from the rack to an appropriate working area. Prepare the anesthesia gas machine by turning on the oxygen flow meter to 1 L/min.
- Place the animal into the induction chamber and close the lid tightly. Turn on the isoflurane vaporizer to 3.5% and monitor the animal until recumbent.
- Remove the animal from the chamber and assess the level of anesthesia by pedal reflex, i.e., firm footpad pinch. Ensure adequate depth of anesthesia before performing the procedure: lack of response to a firm pinch indicates adequate anesthesia.
- Restrain the anesthetized mouse by grasping the loose skin behind the ears with the thumb and index finger of the non-dominant hand. Bulge the eyes by using the index finger to draw back the skin above the eye and the thumb to draw back the skin below the eyes.
- Place the tip of a Pasteur pipette into the eye socket underneath the eyeball (Figure 1, left panel), directing the tip at approximately 45° toward the middle of the eye socket (Figure 1, right panel). Rotate the pipette between fingers during the forward passage. Apply gentle pressure and then release until blood is entering the pipette.
NOTE: Maximum amount of blood that may be withdrawn at one time from this location is about 1% of body weight, e.g., 0.2 mL from a 20 g mouse. - Gently remove the capillary to prevent injury to the eye and place the collected blood in a 1.5 mL centrifuge tube. Close the eyelid and apply mild pressure with gauze to prevent further bleeding. Once fully alert and mobile (usually 3−5 min), return the mouse to its holding cage.
- Allow blood to clot for 30−60 min at room temperature (RT), then centrifuge blood for 10 min at 2,000 x g in a 4 °C refrigerated centrifuge. Using a clean pipette technique, collect serum into a new, labeled 0.5 mL vial. Immediately freeze vial of serum at -80 °C.
- CSF collection with survival procedure
NOTE: This procedure is for survival surgery, and it is based on the protocol published by Liu and Duff in 200815. The mice are anesthetized by a Ketamine (20 mg/mL), xylazine (0.5 mg/mL), and acepromazine (0.5 mg/mL) cocktail administered intraperitoneally.- Move the cages containing mice from the rack to a designated surgery working area. Prepare surgery space in a sterile environment. Ensure that all instruments and materials used are sterilized before surgery.
- Weigh the mouse and calculate the anesthesia volume needed (0.1 mL of anesthesia cocktail for a 20 g mouse). Inject anesthesia intraperitoneally16. After a few minutes, test the mouse by pinching the footpad to ensure adequate anesthesia. If more anesthetic is required, further inject 0.01−0.03 mL of the anesthetic cocktail.
- Use either scissors or a shaver to shave a small area of the head, on the caudal end, medial on the skull, to expose large enough working area for CSF collection. Position the mouse in the prone position on the stereotaxic instrument, and steady the head by using ear bars (Figure 2A).
NOTE: The mouse is laid down so that the head forms a nearly 135° angle with the body (Figure 2A). Once the animal is positioned, a surgical drape is used to maintain a sterile field at the surgical site. Clear adhesive drapes are preferred for CSF collection in mice, as they allow for direct and more focused visualization of the animal. - Swab the surgical site with 30% chlorhexidine diacetate. Using a sterile scalpel, make a sagittal incision of the skin inferior to the occiput to expose muscles overlying the cisterna magna.
- By blunt dissection with forceps, separate the subcutaneous tissue and muscles to expose the cisterna magna (Figure 2B). Use microretractors to hold the muscles apart (Figure 2B) and expose the dura mater meningeal layer over the cisterna magna.
- Gently wash with sterile phosphate-buffered saline (PBS) to remove any possible blood contamination. Blot dry the dura mater with a sterile cotton swab and gently puncture the membrane covering the cisterna magna with a 30 G needle. Quickly and gently insert a small glass capillary tube to collect CSF (Figure 2C).
NOTE: Intracranial pressure allows CSF to flow spontaneously into the capillary (Figure 2C). Depending on the age and size of the mouse, approximately 5−12 µL of CSF is obtained from each mouse. - Carefully remove the capillary tube from the membrane. Connect the tube to a 3 mL syringe through a polyethylene tubing (Table of Materials) and inject the collected CSF into a labeled 0.5 mL tube (Figure 2D). Keep vials in ice.
- Close incision by using polydioxanone suture (PDS) with disposable needle and using buried sutures17. Clean off the area of any dried blood or tissue.
- Inject mice, subcutaneously or intraperitoneally16, with 0.05−0.1 mg/kg of buprenorphine hydrochloride as analgesic treatment. Also, inject subcutaneously 1 mL of sterile saline to prevent dehydration.
- Place the mouse back in a clean and warm cage for recovery. Once the mouse is mobile and able to reach food and water, place the cage back on the rack.
- Centrifuge CSF for 10 min at 1,000 x g in a 4 °C refrigerated centrifuge. Check the degree of blood contamination by visual inspection for identification of xanthochromia and presence of a red pellet in the bottom of the tube. Discard blood-contaminated samples.
NOTE: The formula utilized for the correction of CSF protein amounts in blood-contaminated specimens is based on equation parameters that include protein content in CSF and serum, hematocrit (HCT), and red blood cells (RBC) count in CSF and blood18. However, such a correction strategy cannot be easily applied to mouse CSF specimens due to the small volume, therefore limiting the correction strategy to a visual inspection. - Using a clean pipette technique, collect CSF into a new 0.2 mL tube, leaving behind the pellet with cells. Dilute CSF 1:3 with PBS to reduce volume loss due to aerosol. Immediately freeze the vial of CSF at -80 °C.
- Retro-orbital bleeding procedure for serum collection.
- Serum and CSF collection using non-survival procedures
NOTE: For non-survival fluid collection, CSF collection precedes serum collection as the mouse needs to have a pulse.- CSF collection at necropsy
NOTE: This procedure is for non-survival surgery, and approximately 10−20 µL of CSF is obtained from each mouse. A sterile surgical field is recommended, but not required for non-survival surgery.- Move the cages containing mice from the rack to a comfortable working space. Follow steps 1.1.2.2−1.1.2.7 and 1.1.2.11−1.1.2.12 for CSF collection. Proceed to section 1.2.2 for serum collection.
- Blood collection via intracardiac puncture (open approach)
NOTE: Blood volumes expected is approximately 3% of body weight, e.g., 0.6 mL from a 20 g mouse.- Following CSF collection ensure the mouse is still sufficiently anesthetized by pinching the footpad. If any reaction is observed, administer a second dose of anesthetic. If no reaction is observed, proceed.
- Place the animal on the back and swab skin on the abdomen with 70% alcohol. With surgical scissors, open the thoracic cavity and expose the heart. Insert a 25 G needle (attached to a 3 mL syringe) into the left ventricle and gently apply negative pressure on the syringe plunger. Withdraw needle after blood has been collected.
- Perform a secondary method of euthanasia such as decapitation or cervical dislocation to ensure that the animal is deceased.
- Push the plunger of the syringe down and inject the collected blood into a 1.5 mL vial. Allow blood to clot for 30-60 min at RT and then centrifuge it for 10 min at 2,000 x g in a 4 °C refrigerated centrifuge.
- Using clean pipette technique, collect serum into a new, labeled 0.5 mL vial. Immediately freeze vial of serum at a -80 °C freezer.
- CSF collection at necropsy
2. Protein analysis
- Use a preferred method, e.g., Luminex technology, for quantifying target protein(s) and albumin in matched serum and CSF specimens.
NOTE: Here, an example is given with Luminex magnetic technology, but virtually any technique that measures protein amounts, including enzyme-linked immunosorbent assays (ELISAs), can be applied to the current protocol. Ideally, CSF and serum samples are run for both albumin and target proteins on the same platform. Assay conditions must be optimized for crucial steps in the protocol such as antigen-bead coupling concentration, serum and CSF sample dilutions, best-fit standard curves for each analyte, and buffer composition to reduce non-specific reactivity. If a commercial kit is used for protein(s) measurement, e.g., the immunoglobulin isotyping kit (Table of Materials) used to obtain data presented in Figure 3, manufacturers’ instructions have to be followed.- Upon thawing and prior to analysis, centrifuge CSF and serum samples (2,000 x g for 10 min) and use the supernatant to prevent clogging of the filter plates and/or probe. Follow the assay procedure provided with the kit for appropriate sample dilutions. Otherwise, determine the appropriate dilution for each analyte and fluid. Dilute samples in PBS accordingly.
NOTE: If there are no specific guidance or instructions, dilutions for each analyte and fluid have to be established before the study test, by determining the appropriate dilution ranges necessary to obtain concentration estimates that fall within the most reliable range of a standard curve. Knowing the characteristics of the biological sample to be analyzed, e.g., physiological and pathological concentrations in the fluid, allows trying different dilutions with samples of low, medium, and high analyte content. If the expected range of concentrations in the samples is known a priori, the dilutions can be selected after calculating how many times the sample has to be diluted in order to be within the chosen standard curve range.
CAUTION: By calculating the dilution factors, remember that CSF has already been diluted 1:3. - Prepare a standard curve for each protein of interest, e.g., albumin and IgG as used to generate data in Figure 3, by serial diluting reference standard proteins. During the preparation of standard curves, thoroughly mix each higher concentration before making the next dilution.
NOTE: Regardless of the chosen method of quantification, it is essential to include a standard curve each time the assay is performed to estimate protein(s) concentration in samples. The best choice for a reference standard is a purified, known concentration of the protein of interest. Deciding on the specific dilutions, as well as the number of data points and replicates used to define the standard curve, depends upon the degree of non-linearity in the standard curve. - Select the appropriate antibody-coupled magnetic bead sets (Table of Materials). For individual vials of beads, sonicate each vial for 30 s and vortex for 1 min. Prepare a “working beads mixture” by diluting the bead stocks to a final concentration of 50 beads of each set/µL in assay/wash buffer (PBS, 1% bovine serum albumin [BSA]). Add 50 µL of the mixed beads to each well in a flat-bottom 96-well plate (Table of Materials).
CAUTION: The fluorescent beads are light-sensitive. Therefore, they should be protected from prolonged exposure to light throughout the procedure. - Diagram the placement of backgrounds, standards, and samples on a well map worksheet.
- Add 50 µL of assay/wash buffer to each background well, and 50 µL of each standard to the wells for the standard curve. Load 50 μL of each diluted sample into the appropriate wells last. Wrap the plate with foil and incubate with agitation (~800 rpm) on a plate shaker for 30 min at RT.
- Place the plate on a handheld magnet (Table of Materials) and rest the plate on the magnet for ~60 s to allow complete setting of magnetic beads. Remove well contents by gently decanting the plate and tap plate on absorbent pads to remove residual liquid.
- Wash the plate by removing it from the magnet, by adding 200 µL of assay/wash buffer, by shaking for ~30 s, and finally by reattaching it to the magnet. Repeat washing 3x.
- Dilute the biotinylated detection antibody, i.e., biotin-labeled antibody raised against the protein host species, to 4 μg/mL in assay/wash buffer. Add 50 µL of the diluted detection antibody to each well. Cover the plate and incubate for 30 min at RT on the plate shaker at ~800 rpm. Place the plate on the magnet and repeat steps 2.1.6 and 2.1.7.
- Dilute phycoerythrin (PE)-conjugated streptavidin (SAPE) to 4 μg/mL in assay/wash buffer. Add 50 µL of diluted SAPE to each well. Cover the plate and incubate for 30 min at RT on the plate shaker at ~800 rpm. Place the plate on the magnet and repeat steps 2.1.6 and 2.1.7.
- Remove the plate from the magnet and resuspend the beads in 100 μL of assay/wash buffer. Read wells with a dual laser flow-based detection instrument which allows for the detection of the magnitude of PE fluorescence intensity (FI).
NOTE: The signal, e.g., FI, generated is proportional to the amount of target antigen attached to the surface of the beads. - Export raw data and create standard curves by graphing detection signal FI versus standard protein concentrations. Use the standard curve(s) to calculate the concentration of the analyte(s) in the samples.
NOTE: Albumin is preferentially expressed in g/dL, while proteins of interest are preferentially expressed in mg/dL.
- Upon thawing and prior to analysis, centrifuge CSF and serum samples (2,000 x g for 10 min) and use the supernatant to prevent clogging of the filter plates and/or probe. Follow the assay procedure provided with the kit for appropriate sample dilutions. Otherwise, determine the appropriate dilution for each analyte and fluid. Dilute samples in PBS accordingly.
3. Intrathecal index calculations
- Organize protein concentration values into a spreadsheet and analyze the results by applying the following formulas.
- Calculate Qalbumin:
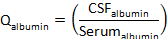
where CSFalbumin and Serumalbumin are concentrations of albumin in matched serum and CSF specimens, respectively. - Calculate Qprotein:
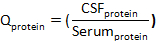
where CSFprotein and Serumprotein are concentrations of target protein(s) in matched serum and CSF specimens, respectively. - Calculate the protein index:
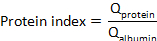
This representative experiment aimed to compare the intrathecal synthesis of IgG in two clinically relevant rodent models of multiple sclerosis (MS): the PLP139-151-induced relapsing experimental autoimmune encephalomyelitis (R-EAE) and the chronic progressive, Theiler’s murine encephalomyelitis virus-induced demyelinating disease (TMEV-IDD). R-EAE is a useful model for understanding relapsing-remitting MS, whereas the TMEV-IDD model features chronic progressive MS19.
For the present study, a quantitative analysis of the intrathecal IgG synthesis in R-EAE (n = 12) and TMEV-IDD (n = 28) has been performed. Both groups were analyzed at the peak of their disease. An additional group of 10 mice was sham-treated and served as age-matched control groups (cR-EAE n = 4, cTMEV-IDD sham n = 6).
A magnetic bead-based approach with the commercially available kit (Table of Materials) was used to measure total IgG in matched serum and CSF specimens. The total IgG value was derived from the sum of the subclass IgG1, IgG2a, IgG2b, and IgG3 values. Albumin was measured with a commercial mouse albumin ELISA kit (Table of Materials) because a Luminex assay for albumin was not available at that time. All measurements were performed carefully following the manufacturers’ instructions. Albumin quotient (Qalbumin) and IgG index (QIgG/Qalbumin) were then used to differentiate blood- versus CNS-derived IgG in the CSF.
As shown in Figure 3A, actual levels of total IgG are significantly increased in the CSF of both rodent models of MS when compared to the corresponding age-matched sham controls (p ≤ 0.026). However, R-EAE mice show significantly enhanced Qalbumin values (p ≤ 0.019), indicating increased permeability of the barrier in these mice (Figure 3B). Conversely, no differences in Qalbumin exist between TMEV-IDD and sham mice (p = 0.49), thus corroborating our previous finding of an intact barrier in TMEV-IDD mice7,8. To further discriminate between transudate and intrathecally produced IgG in R-EAE and TMEV-IDD, the IgG Index was measured, showing significantly higher values in TMEV-IDD mice (p ≤ 0.0006), and therefore intrathecal IgG production in this model (Figure 3C).
An intact barrier in TMEV-IDD mice along with a high IgG index suggests that in this model, antibody is produced within the CNS. Conversely, in R-EAE, a significant barrier breakdown and a low IgG index provide evidence that the CSF IgG is mostly produced by peripheral rather than intrathecal B cells, also suggesting that in this acute model of MS, CSF IgG is mostly derived from serum.
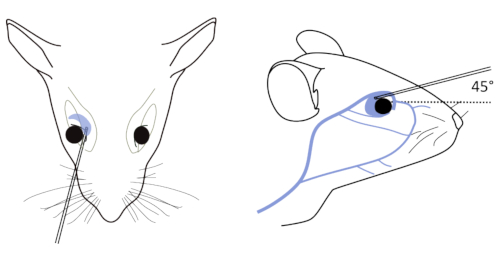
Figure 1: Retro-orbital bleeding of mice. Left: Correct placement of the needle relative to the retro-orbital sinus, the eyeball and the back of the orbit. Right: Pipette location begins in the medial canthus of the eye and glides to the dorsal aspect of orbit. The capillary is inserted at an angle of 45°. Please click here to view a larger version of this figure.
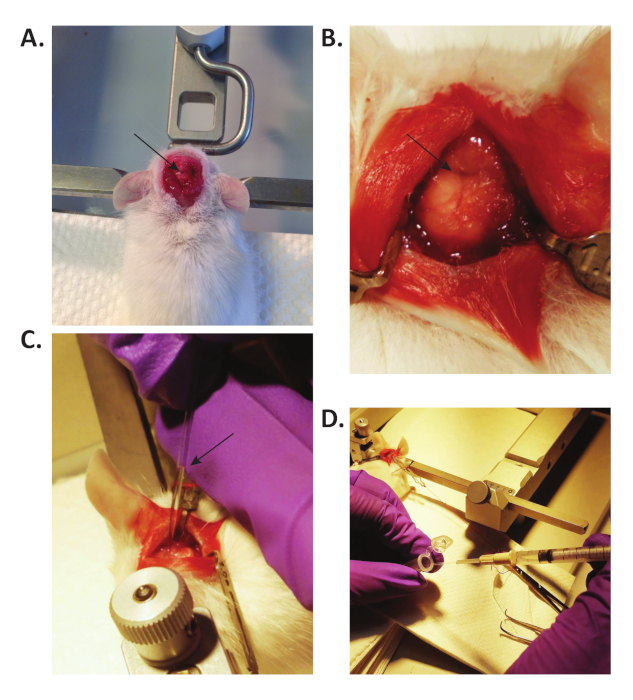
Figure 2: CSF collection in mice. (A) The ear bars support the head of the mouse, and the mouse is laid down so that the head forms a 135° angle with the body. The arrow points to the exposed cisterna magna. (B) By blunt dissection with forceps, the muscles are separated to expose the cisterna magna (pointed by the arrow). Microretractors are used to hold the muscles apart. (C) A small glass capillary tube is used to collect CSF from the cisterna magna. CSF flows spontaneously into the capillary, due to the intracranial pressure. The arrow points to the collected CSF in the capillary. (D) The CSF is transferred into a 0.5 mL tube through a modified 3 mL syringe. Please click here to view a larger version of this figure.
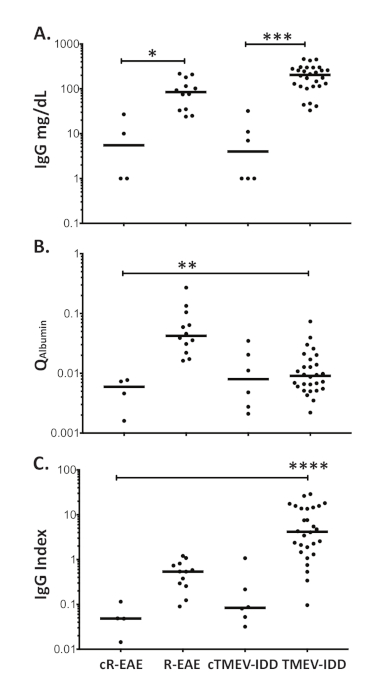
Figure 3: Blood-brain barrier function and intrathecal synthesis of IgG in R-EAE and TMEV-IDD. Dot blot representing (A) the CSF levels of total IgG (mg/dL) measured by Luminex technology, (B) albumin CSF/serum quotients (Qalbumin), and (C) IgG indices (QIg/Qalbumin) in R-EAE and TMEV-IDD mice as well as age-matched control mice (cR-EAE and cTMEV-IDD). Horizontal lines represent the median value for that group. ****p < 0.0001; ***p < 0.001; **p < 0.01; *p < 0.05. Please click here to view a larger version of this figure.
