During classical conditioning experiments, the CS must not induce a spontaneous response in the animals. In the experiments we conducted, only 3% – 4% of the ants performed MaLER in response to the visual cue on the first trial, prior to training. Ants that were undergoing a paired training performed increasingly more MaLER in response to the CS (Figure 3A; logistic regression, N = 51, degrees of freedom (df) = 507, z = 5.949, p < 0.01). The percentage of paired ants that respond to the CS plateaued around 50%, from the third trial onwards. On the contrary, unpaired ants showed no significant increase in MaLER during training (Figure 3B; logistic regression, N = 29, df = 287, z = 0.758, p = 0.45). The occurrence of MaLER in response to the visual cue was significantly higher during paired than unpaired training (logistic regression, N = 80, df = 796, z = -5.306, p < 0.01), which was true for every trial except the first (Table 1).
For examining their short- and a mid-term memory15, the ants were tested either 10 min or 1 h after the last training trial. For both tests, the proportion of ants performing MaLER in response to the CS was significantly higher when they had undergone paired rather than unpaired training (Figure 3C and 3D; Table 1).
During training, individual ants showed substantial variation in the number and type of MaLER that they displayed (Figure 4A and 4B). Only 14% of paired ants responded in every trial from the second or third trial onward, while most ants alternated between trials in which they responded and in which they did not. On those training trials in which ants responded, the degree to which they extended and moved their mouthparts varied. Therefore, we divided MaLER into three different types: FEM, FE, or PE. Typically, ants performed FEM or PE more often than FE. However, only a few ants performed consistently the same type of response; in most cases, ants showed little consistency on the type of MaLER they performed (Figure 4).

Figure 1: Experimental set-up and training scheme. (A) Fix the ant with wax to an insect pin attached to a modeling clay cylinder (orange) and place a custom-made holder underneath it to permit a naturalistic stance. Place it inside a white acrylic box illuminated by two light sources, directly underneath a camera. Use as the conditioned stimulus (CS) a cardboard (blue square) attached to the syringe that delivers the unconditioned stimulus (US), a sugar reward. The inset shows a close-up view of an ant in the holder. (B) The tip of the syringe needle is moved as close as possible to the ant's head but without touching the antennae as shown in this schematic. Train ants through either a (C) paired or (D) unpaired training. This figure has been modified from Fernandes et al.16. Please click here to view a larger version of this figure.

Figure 2: The Maxilla-Labium Extension Response of wood ants. Individual frames from video recordings show ants' mouth part movements during training. (A) This panel shows no response. (B) This panel shows a full extension (FE) of the maxilla-labium that terminates in the glossa. (C) This panel shows a partial extension (PE) with only the maxillary palpus visible. (D) This panel shows a partial extension (PE) of the maxilla-labium structures. (E) Ants undergoing paired training (N = 51) perform a full extension with movement (FEM; dark brown), an FE (mid-brown) or a PE (light brown). This figure has been modified from Fernandes et al.16. Please click here to view a larger version of this figure.

Figure 3: Wood ants form associative memories of a visual cue paired with a sugar reward. (A) The percentage of paired ants (N = 51) performing MaLER in response to the CS presentation significantly increased throughout training. (B) The percentage of ants performing MaLER did not increase significantly throughout unpaired training (N = 29). Ants were tested (C) 10 minutes (paired: N = 15; unpaired: N = 15) or (D) 1 hour (paired: N = 15; unpaired: N = 14) after the last training trial. The percentage of ants responding to the CS during and after paired or unpaired training are represented in brown or grey, respectively. The three types of MaLER are represented in dark (FEM), medium (FE), and light (PE) tones. This figure has been modified from Fernandes et al.16. Please click here to view a larger version of this figure.

Figure 4: Individual performance of ants during training. These panels show the individual performances of (A) paired ants and (B) unpaired ants. The three types of MaLER are represented in dark (FEM), medium (FE), and light (PE) brown or grey. This figure has been modified from Fernandes et al.16. Please click here to view a larger version of this figure.
| Trial | N | df | G (adjusted) | P |
| 1 | 80 | na | na | >0.1 |
| 2 | 80 | 1 | 3.86 | <0.05 |
| 3 | 80 | 1 | 8.41 | <0.01 |
| 4 | 80 | 1 | 6.63 | <0.01 |
| 5 | 80 | 1 | 8.41 | <0.01 |
| 6 | 80 | 1 | 7.5 | <0.01 |
| 7 | 80 | 1 | 10.69 | <0.01 |
| 8 | 80 | 1 | 11.76 | <0.01 |
| 9 | 80 | 1 | 17.13 | <0.01 |
| 10 | 80 | 1 | 17.13 | <0.01 |
| 10 min | 59 | 1 | 5.5 | <0.05 |
| 1 h | 59 | 1 | 4.42 | <0.05 |
Table 1: Comparison of MaLER responses to the CS between ants that had undergone paired and unpaired training, for each trial and test. The number of ants (N), degrees of freedom (df), G-test of independence (G) and p-value are shown. The first trial was analyzed with a Fisher's exact test. This table has been modified from Fernandes et al.16.
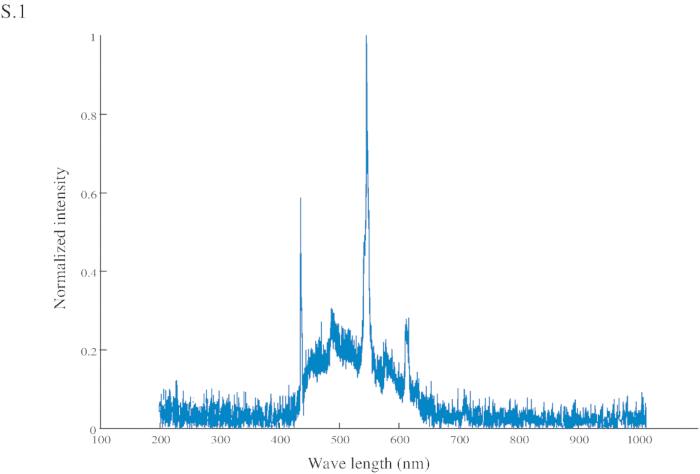
Figure S1: Normalized intensity of the conditional stimuli (CS). The CS has a peak intensity at 545 nm (in the green range) and another at 435 nm (in the blue range).
