In Figure 1, representative images of the scaffold cell seeding and implantation processes are shown. In Figure 1C, note that cells are injected directly into the scaffold. In Figure 1G, note that an incision is made in the back of the mouse, where the subcutaneous pocket is created and the scaffold is implanted. Figure 2 shows the gross morphology of different scaffolds implanted in NSG mice and retrieved after 8 weeks. Note the slight vascularization in hMSC seeded scaffolds (Figure 2A). The co-seeding of human ECs with hMSCs in the scaffold allows for the formation of more relevant vasculature in scaffolds (Figure 2B). Finally, the presence of rhBMP-2 induces bone formation. The retrieved scaffolds are bigger in this case, and they are constituted by bone-resembling hard tissue.
Figure 3 shows the channel configuration setup on the microscope for live-imaging with NDD (details in the figure legend). Figure 4 and Video 1 show human hematopoietic cells in hMSC-coated scaffolds. Scaffolds were explanted 8 weeks post-implantation and after the intravenous inoculation of AF488-hCD45 antibody and 655-VPA. This procedure allows for the visualization of implanted human hematopoietic cells and the vascular structure by two-photon confocal microscopy. In this case, the images show blood vessels (655-VPA) in scaffolds and the long-term engraftment of human hematopoietic cells (AF488-hCD45) in the scaffold parenchyma. Figure 5 and Video 2 correspond to human scaffolds seeded with hECs and hMSCs. 8 weeks after surgery, scaffolds were explanted after the intravenous inoculation of FITC-hCD31 antibody and 655-VPA, and images were acquired with a two-photon confocal microscope, as mentioned before. Images show the participation of hECs in vessel formation in the scaffold, resulting in a murine-human chimeric vasculature.
Figure 6A shows representative data of the approach used to stimulate bone formation in MSC scaffolds. Similar to previous figures, 8 weeks after implantation, the intravenous inoculation of 655-VPA was performed, scaffolds were retrieved, and images were acquired with two-photon confocal microscopy. rhBMP-2-stimulated scaffolds induce the formation of bone tissue, which could be visualized due to the SHG (cyan color in the images) provided by the calcium in the bone. The provided images also show the formation of cavities and vascularized endosteal tissue, which highly resemble the BM endosteal tissue. In Figure 6B and Video 3, hECs were co-implanted with hMSCs. Scaffolds were retrieved after the intravenous inoculation of FITC-hCD31 antibody and 655-VPA, and two-photon confocal microscopy images show the participation of hECs in the neovascularization in a bone-forming scaffold.
Figure 7 shows representative images of histology, a procedure performed to corroborate previously described results. Immunofluorescent images show mouse vasculature, hECs, hMSCs, and long-term engrafted human hematopoietic cells in the scaffold structures. Scaffolds retrieved from mice were fixed and used for immunofluorescence. In the rhBMP-2 carrier bone-forming scaffolds (Figure 7D-F), note the morphology of the tissue, resembling mature bone marrow with adipose tissue. In this bone-forming scaffold, we show that hMSCs are fibroblasts, which would indicate that they contribute to newly formed tissue as stromal cells. We also show human adipocyte marker expression, which would indicate that hMSCs also contribute to adipose tissue formation.
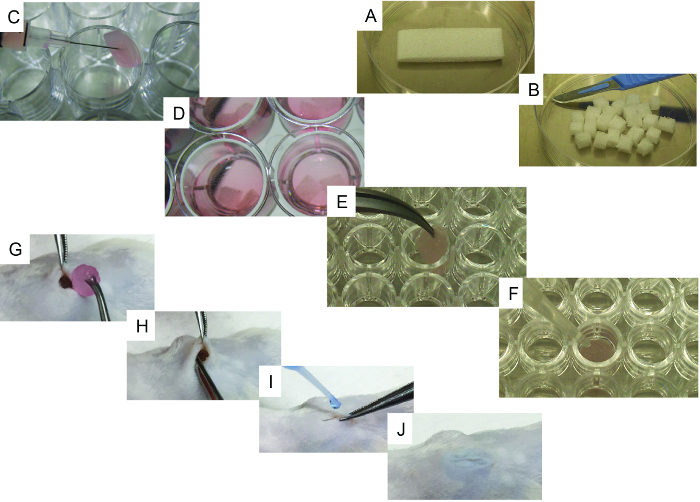
Figure 1. Representative Images of the Cell-seeding and Implantation Processes. A) Initial scaffold and its cutting method using a scalpel. B) 24 pieces obtained from the initial scaffold. C) Scaffold cell-seeding method using a syringe. D) Cell-seeded scaffolds with culture medium, ready to be implanted. E-F) Specific steps for bone-forming scaffolds: E) scaffold being transferred to a 96-well, u-bottom plate and F) representative image of the method used to add rhBMP2, thrombin, and fibrinogen to the scaffold. G-J) Surgical implantation procedure under general anesthesia: G) wound created in the skin and scaffold implantation, H) implantation method, and I-J) wound closing procedure using surgical glue. Please click here to view a larger version of this figure.
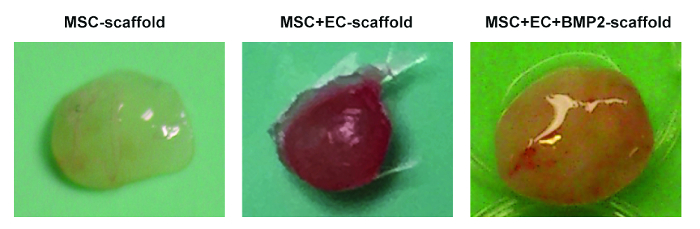
Figure 2. Different Scaffolds Retrieved from Mice. Representative images of MSC scaffolds (left), MSC+EC scaffolds (middle), and MSC+EC+BMP scaffolds (right). Please click here to view a larger version of this figure.
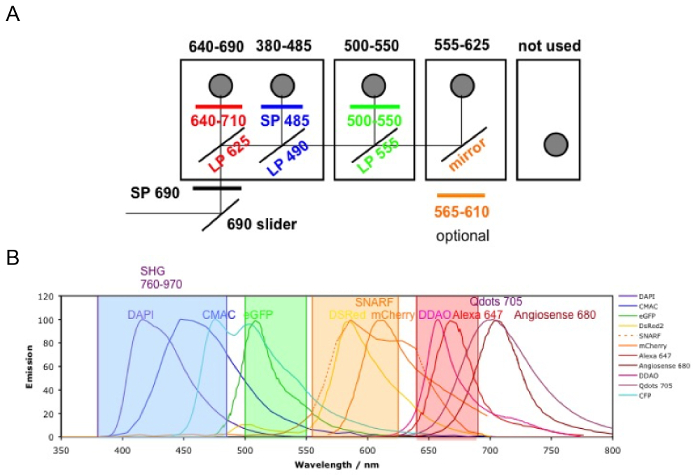
Figure 3. Channel Configuration. The microscope filter setup is shown. A) There are four NDD detector modules: in the first module, there are two filter cubes; the second and third modules have one filter cube each; and the last module has no cube (not used). The first photomultiplier tube (PMT) is for the far-red channel (640 – 690 nm), reflecting the lower wavelengths; the following ones are 380 – 485 nm, 500 – 550 nm, and 555 – 625 nm (the order is always from the lower to higher wavelengths). B) The fluorophore emissions detectable with the above configurations (color coded). Please click here to view a larger version of this figure.
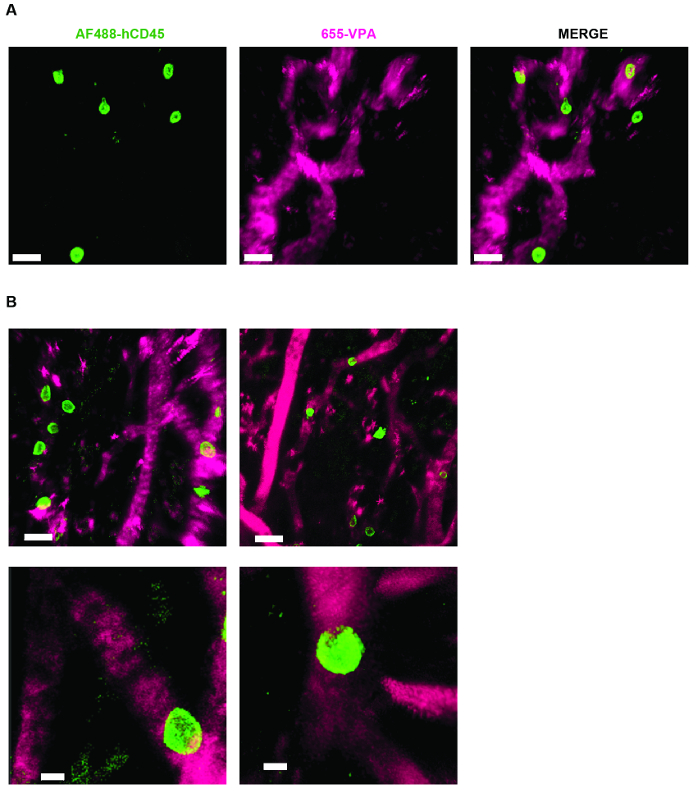
Figure 4. MSC Scaffolds Allow for the Formation of a Niche for Human Hematopoietic Cells. A) and B) 3D reconstructions of Z-stacks taken post-explant, after intravenous inoculation with AF488-hCD45 (green), to label human hematopoietic cells, and 655-VPA (magenta)m to label vasculature. Scale bars represent 20 µm in A and B (upper panels) and 5 µm in B (lower panels). Please click here to view a larger version of this figure.
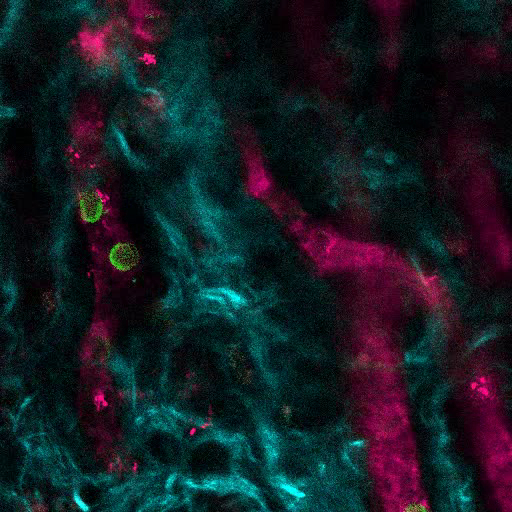
Video 1. MSC Scaffolds Allow for the Formation of a Niche for Human Hematopoietic Cells. 3D reconstruction of human hematopoietic cells (AF488-hCD45) associated with the vasculature (655-VPA) in the MSC scaffold (collagen structures: SHG, cyan). Each stack measures 140 x 140 µm. Please click here to view this video. (Right-click to download.)
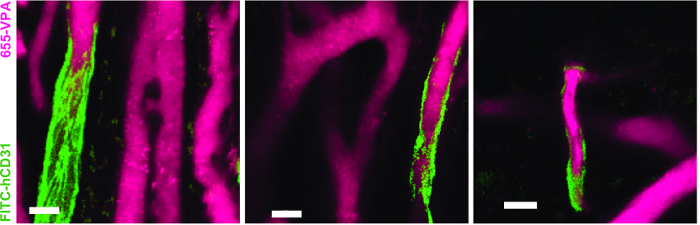
Figure 5. Human ECs Participate in the Formation of Humanized Vessels in MSC Scaffolds. 3D reconstruction of vasculature (655-VPA) lined by ECs of human origin (FITC-hCD31) in MSC+EC scaffolds. Scale bars represent 20 µm. Please click here to view a larger version of this figure.
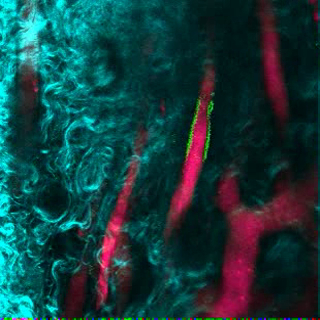
Video 2. Human ECs Participate in the Formation of Humanized Vessels in MSC Scaffolds. 3D reconstruction of human ECs (FITC-hCD31) lining the vasculature (655-VPA) in MSC+EC scaffolds. Each stack measures 240 x 240 µm. Please click here to view this video. (Right-click to download.)
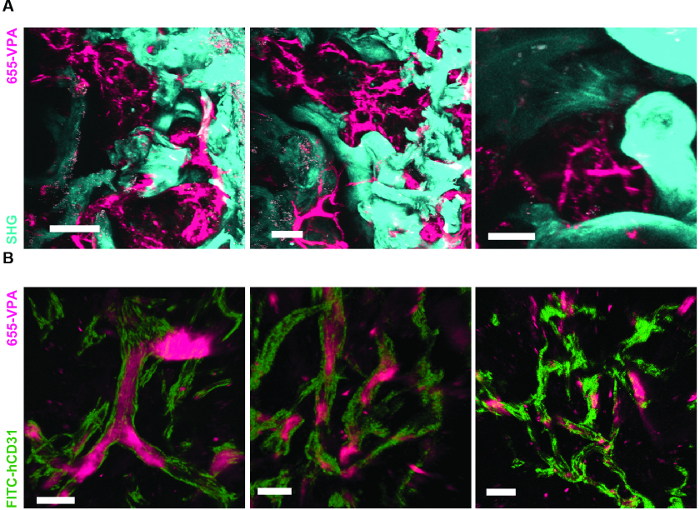
Figure 6. rhBMP-2 Carrier Scaffolds have Bone Surfaces and Humanized Vasculature Similar to the Bone Marrow. A) 3D reconstruction of MSC+BMP scaffolds showing the formation of bone structures (SHG-cyan) and vasculature (655-VPA). Scale bars represent 100 µm (left), 70 µm (middle), and 50 µm (right). B) 3D reconstruction of MSC+EC+BMP scaffolds showing humanized vessels (655-VPA) lined with human ECs (FITC-hCD31). Scale bars represent 50 µm (left) and 30 µm (middle and right). Please click here to view a larger version of this figure.
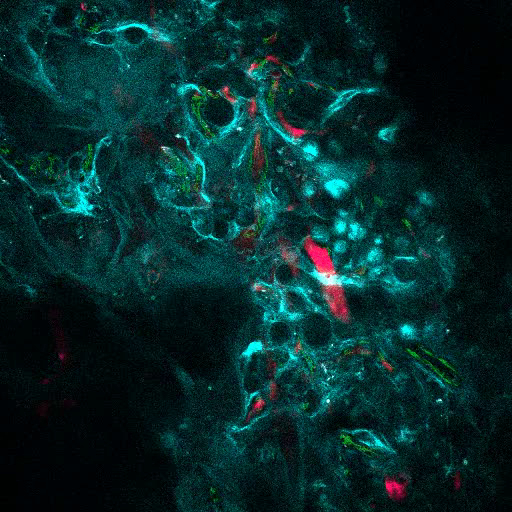
Video 3. rhBMP-2 Carrier Scaffolds have Bone Surfaces and Humanized Vasculature Similar to the Bone Marrow. 3D reconstruction of MSC+VERA+BMP scaffold (bone: SHG; vessels: 655-VPA; human ECs: FITC-hCD31). Each stack measures 600 x 600 µm. Please click here to view this video. (Right-click to download.)
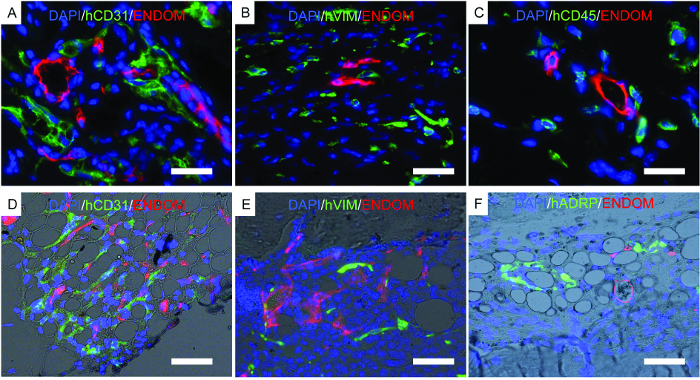
Figure 7. Representative Images of Immunofluorescence Performed on Fixed Scaffolds. A-F) Immunofluorescence studies performed to locate human cells implanted in scaffolds. A-C) Scaffolds implanted with hECs, hMSCs, and hHSCs. D-F) Bone-forming scaffolds implanted with hECs, hMSCs, and hHSCs. The color channels are as follows: A-D) human EC (hCD31) and mouse vascular structure (endomucin, ENDOM), B-E) hMSCs (hVimentin, hVIM) and mouse vascular cells (endomucin, ENDOM), C) human hematopoietic cells (hCD45) and mouse vasculature (endomucin, ENDOM), and F) human adipose differentiation-related protein (hADRP) and mouse vasculature (endomucin, ENDOM). Scale bars represent 10 µm (A and C), 20 µm (B and E), and 40 (D and F) µm. Please click here to view a larger version of this figure.
