Monocyte Purification and Dendritic Cell Differentiation
Monocytes were purified from PBMCs by density centrifugation of peripheral blood (Figure 1A), followed by CD14+ positive selection magnetic separation (Figure 1B) and cultured in complete medium in the presence of GM-CSF and IL-4 to obtain immature dendritic cells (Figure 2A). Addition of vitamin D3 and dexamethasone post GM-CSF and IL-4 resulted in the differentiation of immature moDCs into tolerogenic moDCs (Figure 2B). LPS was added to induce maturation of immature moDCs to mature moDCs (Figure 2C) and tolerogenic moDCs were stimulated with LPS to verify resistance to maturation (Figure 2D). The blood samples used in this work were obtained from healthy donors with previous informed consent.
moDC Characterization
Surface Marker Characterization by Flow Cytometry
Analysis of DC surface markers showed that mature moDCs expressed the highest levels of maturation markers HLA-DR, CD83 and CD86 compared to LPS-treated tolerogenic moDCs, tolerogenic moDCs and immature moDCs (Figure 3). These results demonstrated that tolerogenic moDCs were resistant to maturation as compared to immature moDCs following LPS stimulation. In addition, LPS-treated tolerogenic moDCs and tolerogenic moDCs displayed increased expression of CD14, BDCA3 (CD141) and Immunoglobulin-like transcript (ILT)3 compared to immature and mature moDCs. The tolerogenic moDCs that we generated here is consistent with previous reports 23.
Functional characterization of moDCs
moDCs induced for maturation become immunogenic and release cytokines that promote the proliferation of CD4+ T-cells. We assessed the immunogenicity of the different moDC subtypes by measuring the proliferation of co-cultured T cells. Tolerogenic moDCs were poorly immunogenic compared with mature moDCs, as shown by low alloproliferation of CD4+ T-cells (Figure 4A). Tolerogenic moDCs are characterized by their low IFN-Γ, low IL-12p40, and high IL-10 cytokine production in alloreaction co-cultures with CD4+ T-cells (Figure 4B). Furthermore, increasing the number of tolerogenic moDCs in co-cultures of mature moDCs induced allospecific CD4+ T-cells increased the frequency of CD25high Foxp3+ regulatory T-cells (Figure 4C).
Analysis of Mitochondrial Activity
Red CMXRos is used to reflect the mitochondrial activity to analyze the mitochondrial membrane potential levels in moDCs. Tolerogenic moDCs were observed to have higher mitochondrial activity compared with the other moDC differentiated subtypes (Figure 5A). Next, the rate of mitochondrial oxygen consumption (OCR) is assessed for the different moDC subtypes using a bioanalyzer. OCR measurements allow high resolution insights to the metabolic profile, providing information including but not limited to basal respiration, spare respiratory capacity, proton leak and non-mitochondrial respiration. The measurement of OCR provides a means to assess the ability of cells to respond to stress. The cells are metabolically perturbed by the addition of three different compounds in succession. The first injection is Oligomycin (ATP Coupler) which inhibits complex V of the electron transport chain (ETC), inhibiting ATP synthesis. This step distinguishes the percentage of oxygen consumed for ATP synthesis and the percentage of oxygen consumed to overcome proton leak across the inner mitochondrial membrane. The second injection is FCCP (ETC accelerator) that disrupts ATP synthesis by transporting hydrogen ions across the mitochondrial membrane instead of through the proton channel of Complex V. The collapse of the mitochondrial membrane potential leads to a rapid consumption of energy and oxygen, without the generation of ATP. FCCP treatment can be used to calculate the spare respiratory capacity of cells. Maintenance of spare respiratory capacity under stress conditions is critical to cell survival. This capacity is determined by several factors, including the availability of substrate and functional capacity of the enzymes involved in the ETC. The third injection is a combination of Rotenone, a Complex I inhibitor, and Antimycin A, a Complex III inhibitor. This combination shuts down mitochondrial respiration and OCR is observed to decrease as a result of impaired mitochondrial function (Figure 5C). Tolerogenic moDCs displayed higher basal OCR levels than mature moDCs (Figure 5D). In addition, tolerogenic, LPS-tolerogenic, and immature moDCs showed increased spare respiratory capacity compared with mature moDCs (Figure 5E).
Metabolic Characterization of moDCs
Because lactic acid and protons are released from cells during glycolysis, we analyzed the glycolytic activity of moDCs by performing a real-time analysis of the rate of extracellular acidification (ECAR) (Figure 6B). In the presence of glucose, the glycolytic rate of all moDCs increased compared with the basal stage, with mature moDCs exhibiting higher glycolytic rate than immature moDCs (Figure 6D). Tolerogenic and immature moDCs exhibited higher maximal glycolysis (induced by oligomycin in presence of glucose) compared with LPS-treated moDCs (Figure 6B). The glycolytic capacity of tolerogenic and immature moDCs was higher than mature moDCs (Figure 6E). In contrast to their high glycolytic rate, glycolytic reserve was the lowest in mature moDCs (Figure 6F).
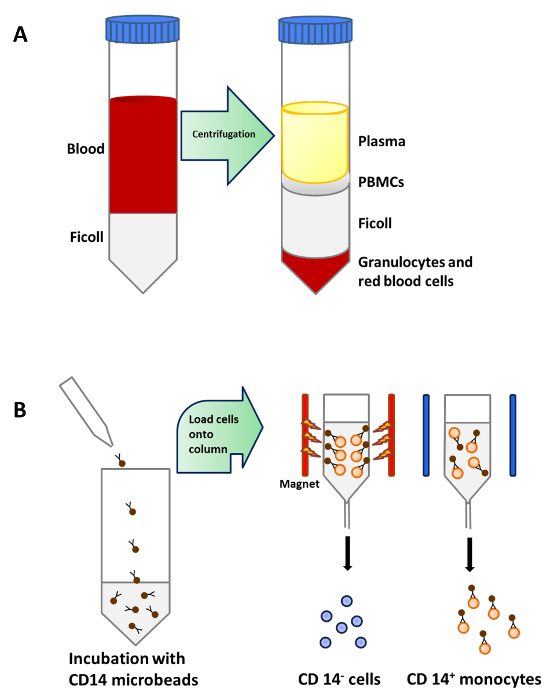
Figure 1: Monocyte Purification from Peripheral Blood. (A) 25 ml of blood is carefully layered onto 15 ml of Ficoll per 50 ml tube before centrifugation. PBMCs are concentrated in a layer below plasma after density centrifugation. (B) PBMCs are incubated with microbeads that are conjugated to monoclonal human CD14 antibodies (isotype: mouse IgG2a) and then loaded onto a column which is placed in the magnetic field of a Separator to isolate CD14+ monocytes. Please click here to view a larger version of this figure.
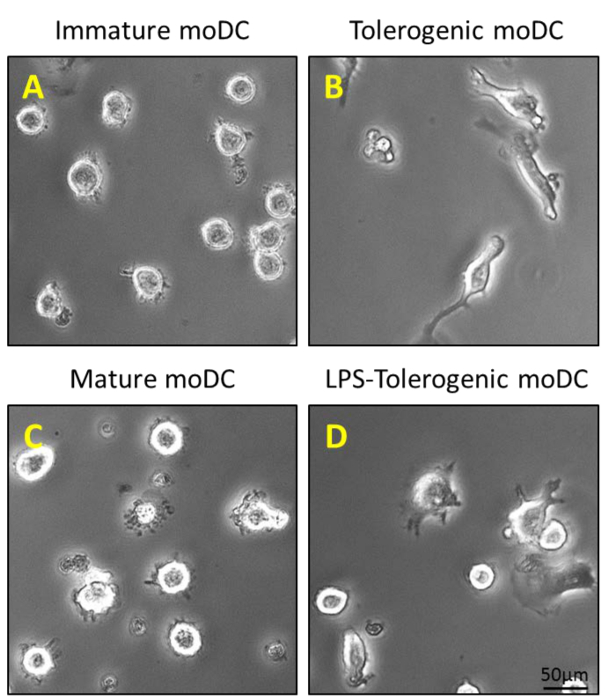
Figure 2: Morphological Characterization of moDCs. (A) 200 ng/ml of GM-CSF and IL-4 is added to purified CD14+ monocytes on Day 0, 4 and 6 to generate immature moDCs; and (B) an additional step of stimulation with 100 nM Vitamin D3 and 10 nM dexamethasone on Day 5 generates tolerogenic moDCs. Immature moDCs and tolerogenic moDCs are stimulated with 1 µg/ml LPS on Day 6 to generate (C) mature moDCs and (D) LPS-tolerogenic moDCs. Please click here to view a larger version of this figure.
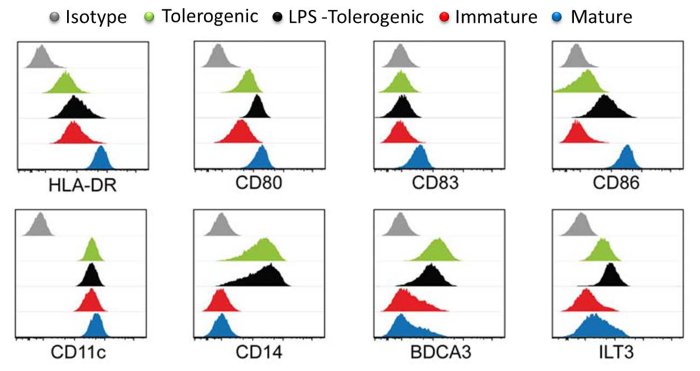
Figure 3: Surface Marker Characterization by Flow Cytometry. Expression levels of surface markers HLA-DR, CD80, CD83, CD86, CD11, CD14, BDCA3 and LT3 in tolerogenic (green), LPS-tolerogenic (black), immature (red), mature (blue) moDCs. Isotype controls are shown in grey. Individual histogram for each cell type is plotted with Y-axis cell count against X-axis log of fluorophore intensity and overlaid. All histograms are representative of four independent experiments. This figure has been modified from J Immunol 194 (11), 5174-5186, doi: 10.4049/jimmunol.1303316 (June 1, 2015). Reproduced and republished with copyright permission. Copyright 2015. The American Association of Immunologists, Inc. Please click here to view a larger version of this figure.
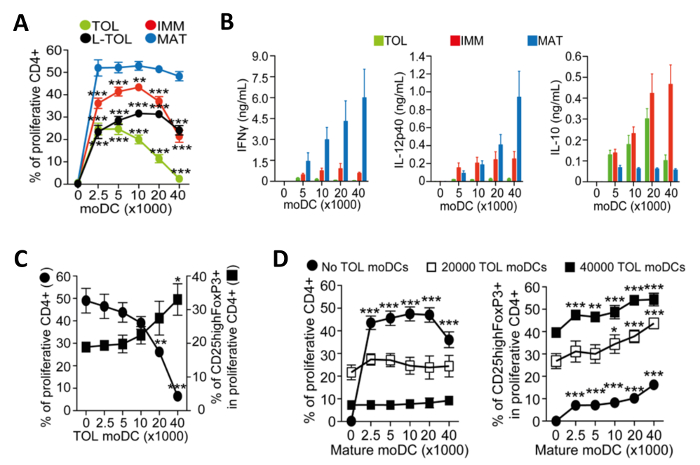
Figure 4: Functional Characterization of moDCs. (A) Quantification of alloproliferation frequency of CD4+ T-cells induced by co-culture with increasing numbers of tolerogenic (TOL; green), LPS-tolerogenic (L-TOL; black), immature (IMM; red) and mature (MAT; blue) moDCs. The data was pooled from four independent experiments; mean + S.E.M. Statistical differences between all moDCs versus mature moDCs were analyzed by two-way ANOVA with Dunnett multiple comparison post-test. (B) Cytokine analysis of IFN-Γ (left panel), IL-12p40 (middle panel) and Il-10 (right panel) in supernatants from alloreactions between CD4+ T-cells co-cultured with increasing numbers of either tolerogenic (green), immature (red) or mature (blue) moDCs. Data were pooled from six independent experiments; mean ± S.E.M. (C) CD4+ T-cell alloproliferation and regulatory T-cells expansion induced by co-culture with mature moDCs in the presence of increasing numbers of tolerogenic moDCs. Left Y-axis, frequency of CD4+ T-cell proliferation. Right Y-axis, frequency of CD25high Foxp3+ cells gated on proliferative CD4+ T-cells. The data were pooled from three independent experiments; mean ± S.E.M. Statistical differences between presence versus absence of tolerogenic moDCs were analyzed by one-way ANOVA with Dunnett multiple comparison post-test. (D) CD4+ T-cell alloproliferation (Left) and frequency of CD25highFoxP3+ cells (Right) induced by co-culture with tolerogenic moDCs in the presence of increasing numbers of mature moDCs. The data were pooled from two to three independent experiments; mean ± S.E.M. Statistical differences between the presence versus absence of mature moDCs were analyzed by two-way ANOVA with Dunnett multiple comparison post-test. This figure has been modified from J Immunol 194 (11), 5174-5186, doi: 10.4049/jimmunol.1303316 (June 1, 2015). Reproduced and republished with copyright permission. Copyright 2015. The American Association of Immunologists, Inc. Please click here to view a larger version of this figure.
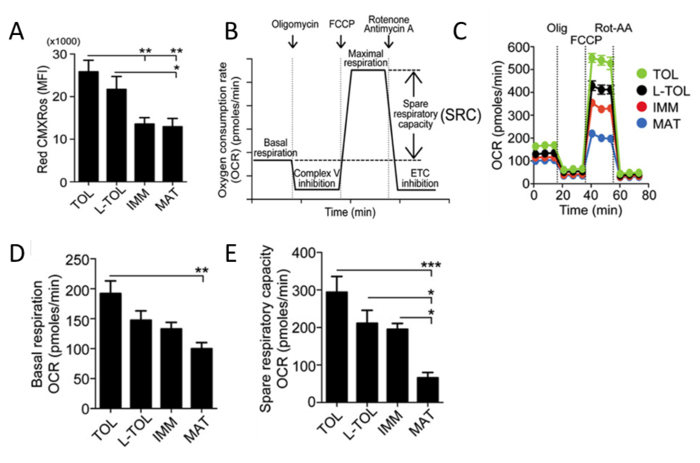
Figure 5: Analysis of Mitochondrial Activity of moDCs. (A) Levels of mitochondrial membrane potential (Red CMXRos) in moDCs were obtained by flow cytometric analysis. The data of four independent experiments were pooled. Mean ± S.E.M. (B) Schematic representation of a real-time mitochondrial respiration. OCR analysis starting from basal respiration and after the addition of oligomycin (complex V inhibition), FCCP (maximal respiration induction), and rotenone/antimycin A mixture (electron transport chain [ETC] inhibition). The mitochondrial SRC (maximal basal subtracted from maximal respiration) is derived from the OCR curve. (C) Representative kinetic study of mitochondria OCR (pmol/min) in tolerogenic (TOL, green), LPS-tolerogenic (L-TOL, black), immature (IMM, red) and mature (MAT, blue) moDCs by using sequential addition of oligomycin (Olig), FCCP, and rotenone/antimycin A (Rot-AA). (D) OCR quantification of basal respiration of moDCs and (E) spare respiratory capacity of moDCs. The data were pooled from five independent experiments. Mean ± S.E.M. This figure has been modified from J Immunol 194 (11), 5174-5186, doi: 10.4049/jimmunol.1303316 (June 1, 2015). Reproduced and republished with copyright permission. Copyright 2015. The American Association of Immunologists, Inc. Please click here to view a larger version of this figure.
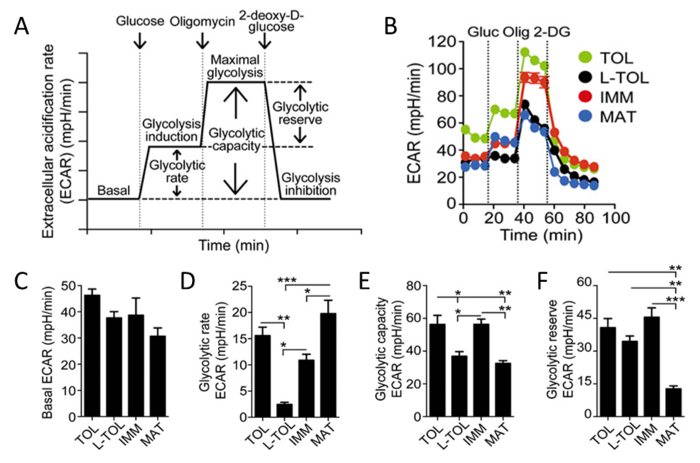
Figure 6: Metabolic Characterization of moDCs. (A) Schematic representation of real-time glycolysis. ECAR analysis starts from basal ECAR, in which the cells were incubated in glucose-free media followed by the addition of glucose (glycolysis induction), oligomycin (which induces maximal cell glycolysis and complex V inhibition), and finally 2-deoxy-D-glucose (glycolysis inhibition). Glycolytic rate (glycolysis induction subtracted for basal ECAR), glycolytic capacity (maximal glycolysis subtracted for basal ECAR), and glycolytic reserve (maximal glycolysis subtracted for glycolysis induction) are derived from the ECAR curve. (B) Representative kinetic study of glycolysis-dependent ECAR (mpH/min) in tolerogenic (TOL, green), LPS-tolerogenic (L-TOL, black), immature (IMM, red), and mature (MAT, blue) moDCs by using sequential addition of glucose (Gluc), oligomycin (Olig), and 2-DG. (C) Bars show basal ECAR levels (D) glycolytic rate (E) glycolytic capacity, and (F) glycolytic reserve of moDCs. The data were pooled from three independent experiments; mean 6 SEM. Statistical differences were analyzed by one-way ANOVA with a Tukey multiple comparison post-test. This figure has been modified from J Immunol 194 (11), 5174-5186, doi: 10.4049/jimmunol.1303316 (June 1, 2015). Reproduced and republished with copyright permission. Copyright 2015. The American Association of Immunologists, Inc. Please click here to view a larger version of this figure.
