The schematic diagram of HE and blood induction from hPSCs by overexpression of transcription factors is shown in Figure 1. ETV2 with GATA1 or GATA2 combination induces pan-myeloid hematopoiesis, while a GATA2, TAL1 +/- LMO2 combination induces predominantly erythro-megakaryocytic hematopoiesis. Both TF combinations directly induced HE cells that subsequently transformed into blood progenitors with a distinct spectrum of hematopoietic differentiation. Differentiation of hPSCs from the pluripotent state to blood producing HE takes on average 4 days. Round blood cells first emerge by day 5 of differentiation. Subsequently, numerous floating blood cells appear and expand in cultures. To maximize the number of floating blood cells, post-transduction cultures can be prolonged to 10-14 days. The desired lineage of cells can be expanded in low-adherent conditions with the appropriate combination of hematopoietic cytokines. Figure 2 demonstrates the induction of blood from hPSCs using ETV2 and GATA1 or GATA2 TFs. Following transduction with ETV2 and GATA2, cells acquire a typical endothelial morphology by day 4 of differentiation, and eventually transform into round blood cells (day 6-8 of differentiation (Figure 2A). Cells collected on days 3-4 of differentiation show a typical VE-cadherin+CD226+CD73– HE phenotype (Figure 2B, 3B). By day 7 of differentiation more than 50% of VE-cadherin+ cells acquire the hematopoietic CD43+ phenotype (Figure 2C). CFC-assays of GATA2/ETV2-induced cells reveal that colonies are comprised of high proliferative potential (HPP) CFCs, Erythroid (E) CFCs, macrophages (M) CFCs, and granulocytes (G) CFCs (Figure 2E). The hematoendothelial differentiation in hPSCs following overexpression of GATA2 and TAL1 alone or with LMO2 is shown in Figure 3. HE stage is best identified in cultures with GATA2 and TAL1. The addition of LMO2 accelerates differentiation and markedly contracts HE stage. The GATA2 and TAL1 combination alone or with LMO2 demonstrates differentiation and maturation of blood progenitors to mostly erythroid and megakaryocytic cells Figure 3D and 3E. Figure 4 shows optimization of transduction efficiency in H1 hESCs using GFP lentivirus (A) and cell death and differentiation efficiency in cultures transduced with GATA2/TAL1/LMO2 at high and low MOI.
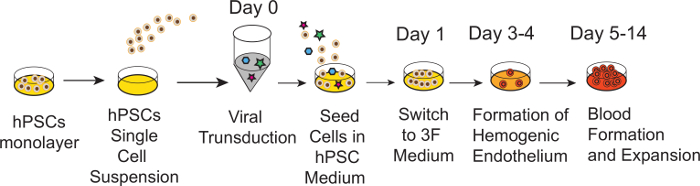
Figure 1: Schematic diagram of protocol for induction of hemogenic endothelium and blood cells via the forced expression of TFs. Cartoon outlines major experimental steps and timelines. Following preparation of single cell suspension, hPSCs are transduced with TFs and seeded on matrix in complete commercial serum-free medium. Next day, medium is replaced with growth factor free commercial medium supplemented with bFGF, TPO and SCF (3F Medium). Following 3-4 days of culture, HE is formed. Subsequently, HE undergoes endothelial to hematopoietic transition with formation of multiple blood cells.
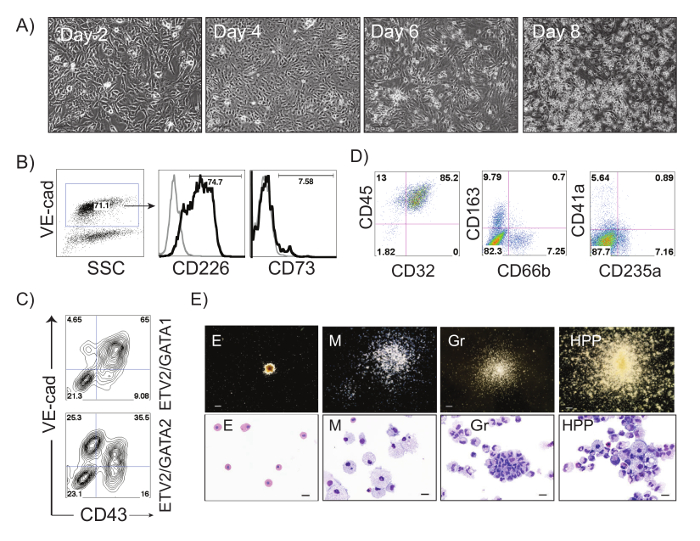
Figure 2: ETV2/GATA2-induced differentiation of hPSCs. (A) Phase contrast images of ETV2/GATA2-induced differentiation in H1 hESCs at days 2, 4, 6 and 8 after transduction; Scale bar, 100 µm. (B) Flow cytometric analysis of ETV2/GATA2-induced cells undergoing endothelial to hematopoietic transition at day 3 of differentiation: VE-cadherin+ HE cells express CD226 and lack expression of CD73. A small percentage of endothelial cells express CD73 (non-HE). (C) Flow cytometric analysis of ETV2/GATA1- and ETV2/GATA2-induced H1 hPSCs on day 7 post-transduction. (D) Flow cytometric analysis of ETV2/GATA2-induced hematopoietic cells expanded in medium supplemented with FBS and SCF, IL3, IL6, GM-CSF, G-CSF and EPO for 14 days. (E) Types of hematopoietic colonies formed from ETV2/GATA2 transduced cells in CFC-assay and corresponding Wright-stained cytospins representing Erythroid (E), Macrophages (M), Granulocytes (Gr) and High Proliferative Potential colonies (HPPs). Scale bars for CFC-assay, 250 µm, for cytospins, 20 µm. Please click here to view a larger version of this figure.
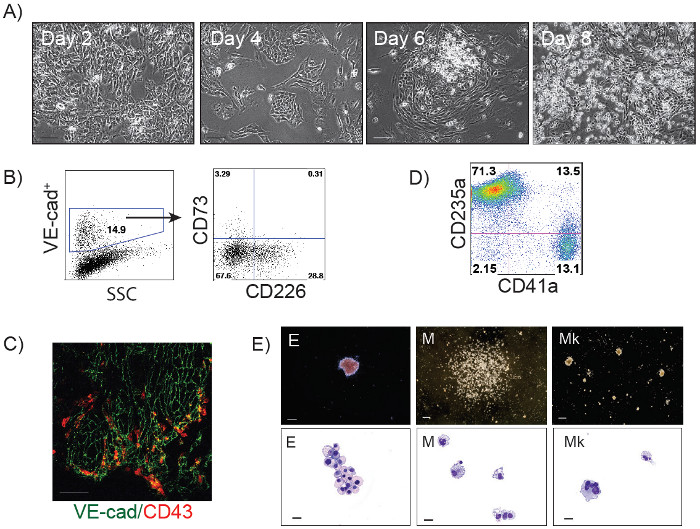
Figure 3: GATA2/TAL1 and GATA2/TAL1/LMO2-induced differentiation of hPSCs. (A) Phase contrast images of GATA2/TAL1/LMO2-induced differentiation in H1 hESCs at days 2, 4, 6 and 8; Scale bar, 100 µm. (B) Flow cytometric analysis of GATA2/TAL1-induced cells undergoing endothelial to hematopoietic transition on day 3 of differentiation: VE-cadherin+ cells acquire expression of HE marker CD226. A small percentage of endothelial cells express CD73 (non-HE). (C) Immunofluorescent staining of GATA2/TAL1-induced differentiation in H1 hESC, day 5; VE-cadherin+ cells (green) acquire expression of hematopoietic marker CD43 (red). (D) Flow cytometric analysis of GATA2/TAL1/LMO2-induced hematopoietic cultures in suspension on day 14 post-transduction following expansion in serum-free medium supplemented with SCF, TPO, EPO and bFGF; (E) Types of hematopoietic colonies formed in CFC-assay by GATA2/TAL1/LMO2 differentiated cellsand corresponding Wright-stained cytospins representing Erythroid (E), Macrophages (M), and Megakaryocytes (Mk). Scale bars for CFC-assay, 250 µm, for cytospins, 20 µm. Please click here to view a larger version of this figure.
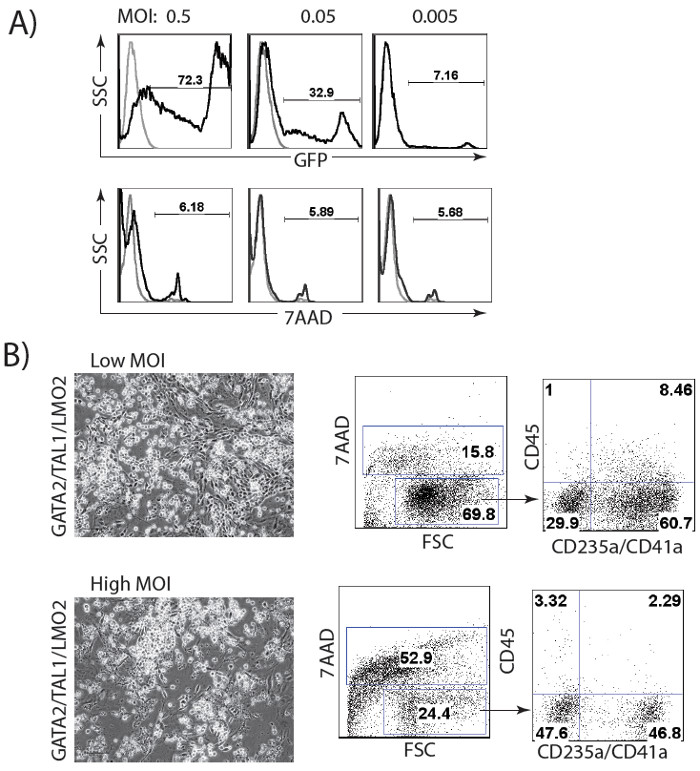
Figure 4: Optimization of lentiviral transduction procedure. (A) Transduction of H1 hESCs with different MOI of GFP lentivirus. (B) Cell viability and differentiation efficiency in cultures transduced with GATA2/TAL1/LMO2 at low (1 for each virus) and high (2 for each virus) MOI.
