The study describes a new method to observe the temperature preference in mosquitoes. In this method, mosquitoes are released into a tube that is connected to two incubators with independently controllable temperatures. In this manner, the mosquitoes are allowed to freely choose between two temperatures without disrupting their natural behaviors and mechanism of expressing this choice (e.g., flying).
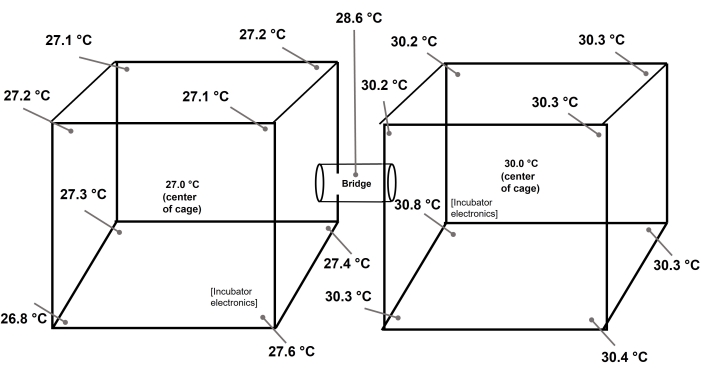
Our first representative experiment used the mosquito optimal temperature of 27 °C in both chambers. During the repetitions of this experiment, mosquitoes were observed to be freely flying between both cages for the entire 30 min, and in all replicates, there were near equal numbers in each of the two chambers. This confirmed the experimental intention of allowing the mosquitoes the ability to freely choose between cages while exhibiting their natural behaviors (flying). Conversely, the second representative experiment utilized the attractive optimal temperature of 27 °C in one chamber and a sub-optimal and hence repelling temperature of 30 °C in the second chamber. As expected, mosquitoes consistently selected the optimal temperature chamber at high significance, even when we swapped the incubators to avoid bias.
We also tested the setup for a different insect, D. melanogaster (fruit flies), representing another ectotherm model organism. One chamber was set to the optimum temperature of D. melanogaster, 25 °C, and the other was set to 3 °C higher, 28 °C. Similar to mosquitoes, fruit flies also favored their optimal temperature and avoided the warmer chamber. This demonstrates that the protocol is suitable for a range of ectotherms.
Description of critical steps in the protocol
The main critical step in the protocol is insect handling, as it generates the possibility of insects escaping. This can be prevented by determining that there are no holes large enough for escape in the cages used, that the rubber bands/cable ties used to secure the mesh sleeves to the bridge are tight, and that the cover for the insect insertion hole on the bridge is securely attached and sealed.
It is also crucial to ensure insects do not escape before or after the experiment, particularly when the insects are required for downstream experimentation or later time points for various temperature choices. This can be done by anesthetizing the insects before placing them in the acrylic bridge (using ice for Drosophila and CO2 for mosquitoes) and releasing CO2 into the bridge to knock down the insects after the experiments, prior to calculating. The usage of CO2 is ideal for mosquitoes since it won't affect the behavioral results21. In flies, exposure to CO2 can alter their flying behaviour23, hence it is recommended to use ice22.
Counting of insects is also a critical step, to ensure the numbers of insects are equal before and after the experiment for accurate results. In order to do this, we recommend the usage of a CO2 pen once the experiment is completed to knock down the insects that are located in the bridge. This will help move the insects to either side of the chamber, hence reducing the number of escapees. We also highlight in the protocol that insects can be caught in the sleeves of the cages during cage separation; therefore, ensure these are checked thoroughly during counting.
Potential modifications and troubleshooting of the technique
The main difficulty with this technique is the flexible mesh of the cage sleeves resulting in gaps or hiding places and hence insect escape or trapping. There are some potential modifications, if needed, to improve the technique. We suggest using two or more rubber bands to ensure that the bridge is secured properly in between the chambers without leaving any potential space for the insects (loose mesh creates a hiding space for insects). We also advise particular care to pull the mesh sleeve taut, as described in step 2.6, when assembling the apparatus.
Small form factor incubators are usually heated only (i.e., have no active cooling), as was the case for the incubators used here. Consequently, using temperatures around or below the ambient room temperature will require the experiment to be performed in a cold room to ensure that the temperatures set for the incubators will go as low as desired.
In addition, this setup can also be used for BSL 3, where a class three biosafety cabinet (glove box) is needed. In this case, the glove box needs to be big enough to fit the entire apparatus. The experiment described in this protocol is ideal for experiments in a glove box because everything required will be contained within the glovebox and, importantly, the possibility of insects escaping is minimal.
Finally, there is enough space in the incubators to add external light or a humidity source without affecting the insects in the cages. Depending on the insect species or experimental design, an LED lamp with 1 cm thickness can be easily placed on top of the cage inside one or both incubators. Providing light to both and offering a temperature choice can be a more realistic protocol for some photosensitive experimental designs, or only providing light (or humidity) to one chamber is a possible modification to the protocol to assess light/humidity choice.
Advantages of this technique in the context of dual choice temperature preference assays
The method described here presents an alternative to the traditional temperature gradient method described in previous studies10,13,14,16. In most of these studies, a large horizontal aluminum block with a thermal gradient is used, while the mechanism of generating this gradient varies, including heating/cooling blocks, water baths, etc. In these instances, the temperature gradient is produced on the surface of the aluminum block (rather than the air temperature in a cage). Consequently, most (but not all) alternative techniques do restrict the flight ability of insects more than this protocol. Here, insects can fly relatively freely between cages, allowing for a more realistic expression of natural behaviors in choice. It would even be possible to scale-up this experimental apparatus using larger cages and incubators, for example, for larger insects.
In addition to the natural behavior advantage, we also demonstrate very high temperature uniformity within the two chambers, enabling simple scoring and a clear selection of two large single temperature chambers. The use of a binary large chamber design such as this may reduce noise in the data, where, for example on a gradient apparatus, any incidental movement of the insects will alter position on the gradient and hence their perceived temperature preference.
The technique described here is also very simple and low cost. This technique does not need extra appliances to set the temperatures (i.e., a water bath10 and/or a hot plate11,12,13,14,15), no specialist equipment besides a cut acrylic tube and drilled holes, and no camera18,19 or sophisticated software19 for analysis. Such components used in other techniques can be expensive and/or require significant expertise and testing to begin experiments.
This technique can also be replicated with different devices that use batteries if there is no external power supply, making the system ideal to conduct experiments in the field. Furthermore, the same apparatus could be slightly modified to study other binary choice preference situations, such as light versus dark, high/low humidity, etc., either in the laboratory or field.
The full-sized apparatus in the protocol is significantly smaller than temperature gradient setups, allowing for an easier fit inside a BSL 3 glovebox as described above. Further, the insects are easier to contain, as they can be knocked down with CO2 at the end of the experiment, and the cages can be quickly resealed after separation from the bridge. These containment advantages are ideal for BSL 3 work.
We do however acknowledge that our apparatus only allows for a binary decision rather than a free choice along a gradient, which, depending on the application, may require additional runs to identify optimal temperatures.