SSM-based electrophysiology has been extensively used for the characterization of electrogenic transporters. In the protocol presented here, we show how to use SSM-based electrophysiology to classify nanobodies targeting a secondary transporter (here a bacterial choline symporter) based on their inhibitory and non-inhibitory properties. One of the most useful features of this technique is that it allows for the high-throughput screening of multiple buffer conditions. This particular characteristic is beneficial for the analysis of nanobody libraries, which after the selection of binders can be constituted from a few to dozens of nanobodies. In a standard experiment, a stable lipid monolayer is assembled on a sensor chip. After applying the proteoliposomes preparation containing the choline transporter, a check for good conductivity and capacitance is performed as this is essential for the success of the experiment. In case that the integrity of the membrane is compromised during an experiment, which is easily observed due to the high noise background currents, changing to a new chip is recommended as recovering low noise conditions is rather difficult. In general, we have observed very good reproducibility among measurements of transport and inhibition by nanobodies when using different chips.
To decide about the substrate concentration to be used during a screening of nanobodies, electrogenic transport was first measured under different substrate concentrations to determine EC50 (Figure 1B,C). A substrate concentration that corresponds to saturating conditions was selected (Figure 1C). This substrate concentration was then kept constant in all activating buffers. For this particular example, we selected 5 mM choline.
For the screening of nanobodies, the nanobody must be added to both non-activating and activating buffers. When nanobodies were added to only the activating buffer, it was not possible to observe inhibition of the electrogenic transport. We speculate that this is due to an incomplete occupation of all nanobody binding sites in the transporter population on the chip, thereby revealing the importance of pre-incubation with nanobodies in non-activating conditions. To ensure that all sites are likely to be occupied, a time delay step was included during the application of the first non-activating buffer step to allow the saturation of nanobody binding sites on the transporter population. Incubation times ranging from 2-60 min have been tested with reproducible results. Keep in mind that optimal times of incubation depend on the nature of the nanobody binder and its concentration during the experiment (as well as the concentration of transporter in proteoliposomes on the chip). Therefore, it is recommended to try different incubation times. In any case, as a rule of thumb, the lower the nanobody concentration, the longer the incubation time required. We tested incubation times of 2 min, 20 min, 30 min, and 60 min for different nanobodies but did not detect further transport inhibition.
The effect of inhibitory nanobodies on electrogenic transport is visualized from the decrease of peak currents amplitudes (Figure 2A,C,D). Non-inhibitory nanobodies, on the other hand, do not affect peak currents. After running the washing protocol to allow nanobodies unbinding, a recovery of 80 to 95% of the initial peak current amplitude was observed (Figure 2A,C,D). We have performed a similar experiment but in the presence of liposomes without the transporter protein. When changing from non-activating to activating conditions, no significant artifact currents was introduced by nanobodies present in these buffers (Figure 2B). Running this control experiment is recommended as it is important to know whether changes in peak currents arise from artifacts or not.
After the selection of nanobodies with inhibitory properties, we determined IC50 values for individual nanobodies (Figure 3A,B). For this particular experiment, it is recommended to start with a low concentration of nanobodies and then move towards high concentration during the assay. The calculation of the inhibition for each concentration was then performed by comparing peak currents measured before and after the application of nanobody. To avoid unspecific binding of nanobodies to surfaces, which can be particularly problematic when using low nanobody concentrations, it is advised to follow a similar protocol to that described by Kermani et al.37, where 50 µg/mL of bovine serum albumin was added to the buffers, preventing this deleterious effect. Adding detergents such as Tween or Triton for this purpose should be avoided as these would dissolve lipid membranes.
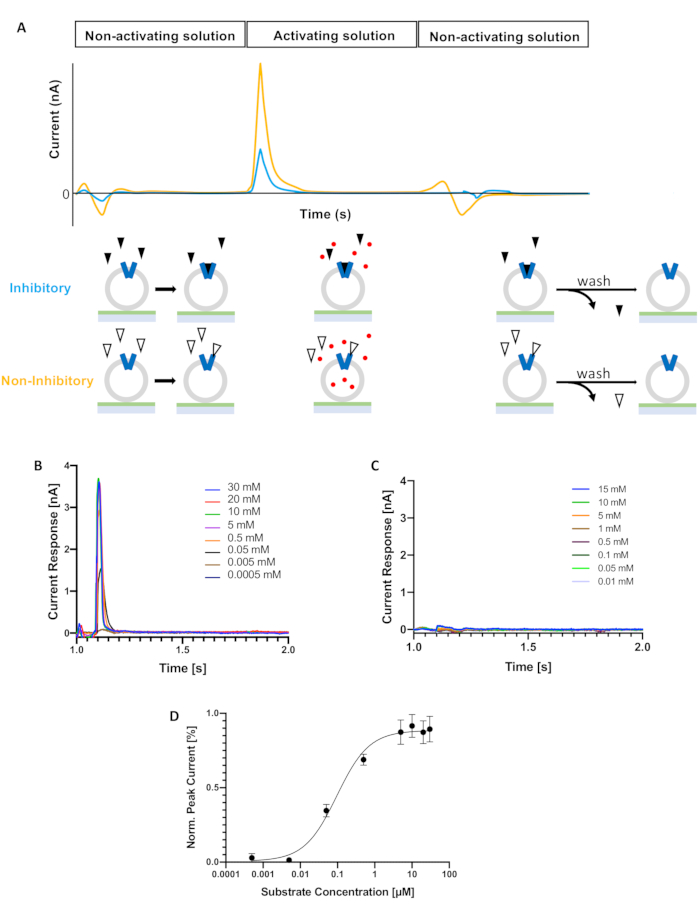
Figure 1: SSM-based electrophysiology. (A) Protocol for transient currents measurement. A non-activating solution is replaced by an activating solution followed by the flow of non-activating solution to restore initial conditions. During the first step, nanobodies bind to the transporter. When switching to the activating solution, the substrate gradient drives the electrogenic transport (orange curve). In the presence of an inhibitory nanobody, the peak current shows a smaller amplitude (blue curve). After finishing the protocol and running solutions without nanobody (wash), unbinding of nanobodies occurs. In the schematic, proteoliposomes with reconstituted protein (blue) are immobilized on the SSM sensor. Triangles and red circles represent nanobodies and substrate, respectively. (B) Electrogenic choline transport in the absence of nanobodies. Peak currents measured during activating conditions are shown for different substrate concentrations. (C) Representative measurement of currents during activating conditions in the absence of transporter protein at different substrate concentrations. (D) Plot of substrate concentration versus peak currents amplitude. The EC50 determined was 95 ± 11 µM choline. Error bars indicate standard deviation (n=3 biological replicates, n=3 technical replicates). Please click here to view a larger version of this figure.
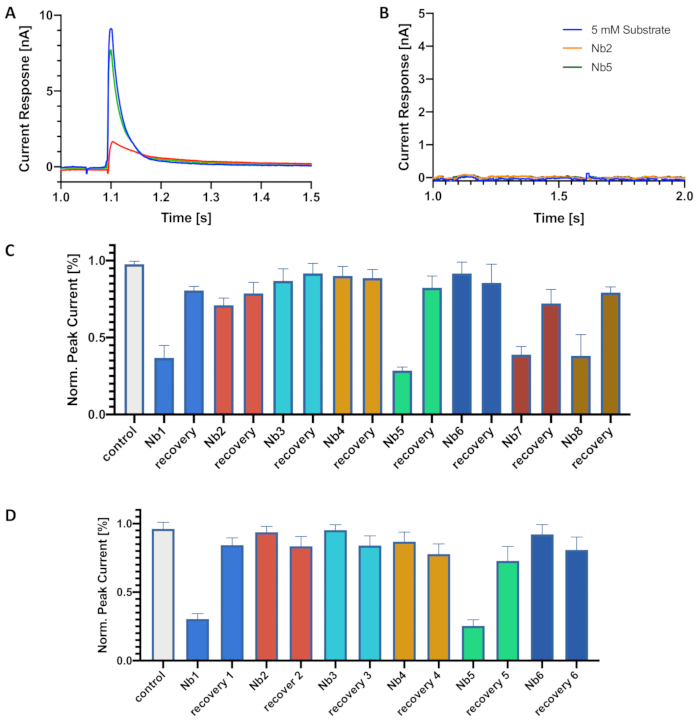
Figure 2: Screening and classification of inhibitory and non-inhibitory nanobodies. (A) Electrogenic choline transport in the presence of a nanobody. Peak currents measured during activating conditions are shown in the absence of nanobody (blue), in the presence of an inhibitory nanobody (red), and after nanobody unbinding (green). (B) Measurement of currents during activating conditions in the absence of transporter protein. Traces show recordings in the absence of nanobody (blue), in the presence of an inhibitory nanobody (green), and in the presence of a non-inhibitory nanobody (red). (C,D). Histograms showing peak currents measured during activating conditions in the presence of nanobodies and after nanobody unbinding (recovery). Panel C shows the results of measurements using individual chips per nanobody. Panel D shows the results from a serial measurement using one chip. Nanobodies are indicated as Nb. Error bars indicate the standard deviation (n=3 biological replicates, n=2 technical replicates). Please click here to view a larger version of this figure.
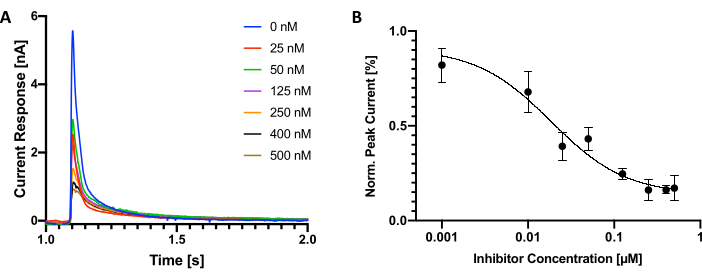
Figure 3: Determination of IC50 of an inhibitory nanobody. (A) Electrogenic choline transport and inhibition by a nanobody. Peak currents measured during activating conditions are shown for different nanobody concentrations. (B) Plot of peak currents amplitude vs nanobody concentration from a serial measurement with an inhibitory nanobody. The IC50 determined was 18 ± 2 nM. Error bars indicate the standard deviation (n=3 biological replicates, n=3 technical replicates). Please click here to view a larger version of this figure.
