24 hours after plating, day 1, iPSCs should be 50%-90% confluent. On day 2, DE should be 90%-95% confluent. During DE induction, it is common to observe significant cell death on day 4 but attached cells will retain a compact cobblestone morphology (Figure 2b). Discontinue differentiation if the majority of adherent cells detach and consider shortening exposure to DE media with activin A by 6-12 h. During AFE induction, cell death is minimal, and cells remain adherent, but will appear smaller and more heterogeneous. Passaging the cells on day 7 must only be done if the yield of double positive SOX2 and FOXA2 is >80%. After passaging into basement membrane matrix for 3D LPC induction, small spheroids will first appear, then grow and some may begin to branch. Gene expression profiles for successful endodermal differentiation include increased SOX17 at DE, increased FOXA2 and SOX2 with decreasing SOX17 and the first appearance of NKX2-1, and increased NKX2-1, along with the presence of SOX2 and FOXA2. Consistent with early embryonic development, the ventralization of AFE occurs for lung bud development (NKX2-1+) and dorsalization of AFE occurs for gastrointestinal development (SOX2+). Cultures at LPC will have a mix of both lung and gastric progenitors.
Lung organoid induction from LPC has been performed using various methods. Some groups sort the cells using NKX2-1 fluorescent reporters or a surface antigen proxy (CPM, CD26lowCD47high). But those lung organoids contain alveolar type II like cells without alveolar type I cells or mesenchyme. Other groups have collected cell clumps that bud off the AFE/LPC monolayer and embedded them into basement membrane matrix. Those organoids contain a mixed population of lung epithelial and mesenchymal cells but take months to culture19. Our protocol includes both the presence of epithelial and mesenchymal cells. The WLOs express proximal epithelial cell markers p63 and KRT5 (basal cells) and SCGB3A2 (club cells) as well as distal epithelial cell markers HOPX (ATI) and proSPC, SPB, and NKX2-1 (ATII). They also express the mesenchymal marker Vimentin at the LPC stage, as well as in the whole lung organoids. PDGFRα is a marker for fibroblasts that have an important function in the lung during sacculation and alveolarization20 and is co-expressed with the transcription factor important in distal cell differentiation, SOX9 (Figure 3).
Our method efficiently generates NKX2-1-expressing LPC 3D cultures using signaling molecules that occur in fetal lung development to form early lung organoids. When passaging LPCs into GFR-basement membrane matrix medium for lung organoid induction, it is imperative not to over-dissociate into a single cell suspension, but to instead to retain small clumps of cells (10 cells/clump). Cell counting will not be completely accurate, but still necessary to avoid over confluence during the 3-week lung organoid differentiation.
Lung organoid induction should yield small, branching organoids by day 6 of induction (day 23 of differentiation). These should continue to grow during the organoid branching step and maturation step. Twenty-four hours after the introduction of dexamethasone, cAMP, and IBMX, the branches should expand into transparent spheres. Whole lung organoid analysis can be performed at the end of the differentiation, or the WLOs can be passage into fresh basement membrane matrix with GFR or cryopreserved by freezing down in 10% DMSO.
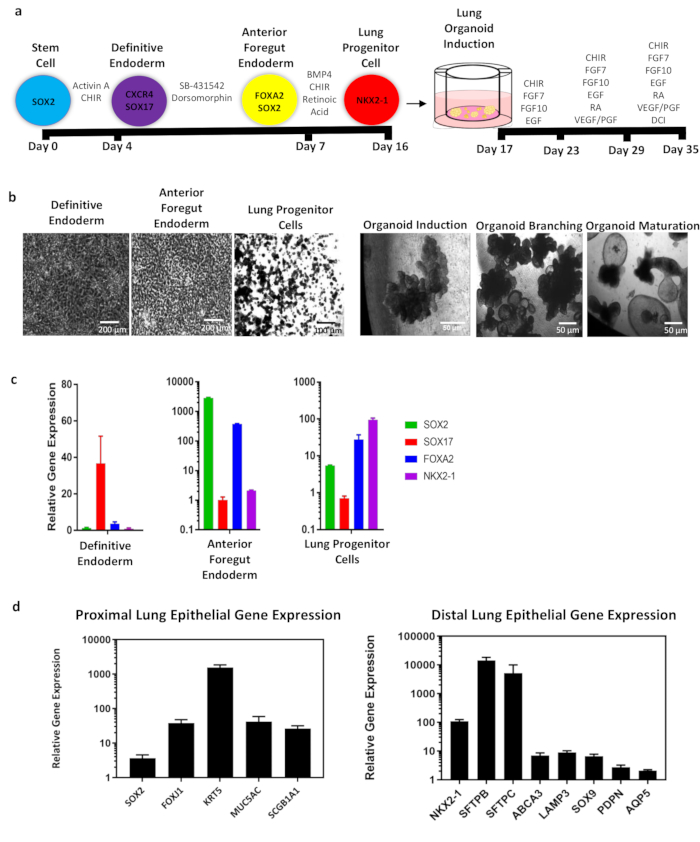
Figure 1: Overall schematic of whole lung organoid (WLO) differentiation from hiPSCs and representative data. (a) Schematic of WLO differentiation from hiPSCs. Circles represent endodermal cell type with identifying markers. Timeline of differentiation is indicated in black bars. Growth factors and/or small molecules for induction of endodermal and lung organoid populations. In summary, stem cells are differentiated into definitive endoderm, anterior foregut endoderm and into lung progenitor cells in approximately 16 days. These cells are then passage into GFR-basement membrane matrix medium containing medium inserts and undergo lung organoid induction, branching, and maturation. The total differentiation takes approximately 35 days. (b) Representative phase contrast images of the cells at major endodermal stages and 3D images of whole lung organoids. Scale bar size as indicated in panel. (c) qRT-PCR analysis of lung development markers during endoderm and (d) whole lung organoid differentiation of proximal and distal cell markers. All data represents an average of 3-5 biological replicates. Error bars represent standard error of the mean and are normalized to actin. Please click here to view a larger version of this figure.
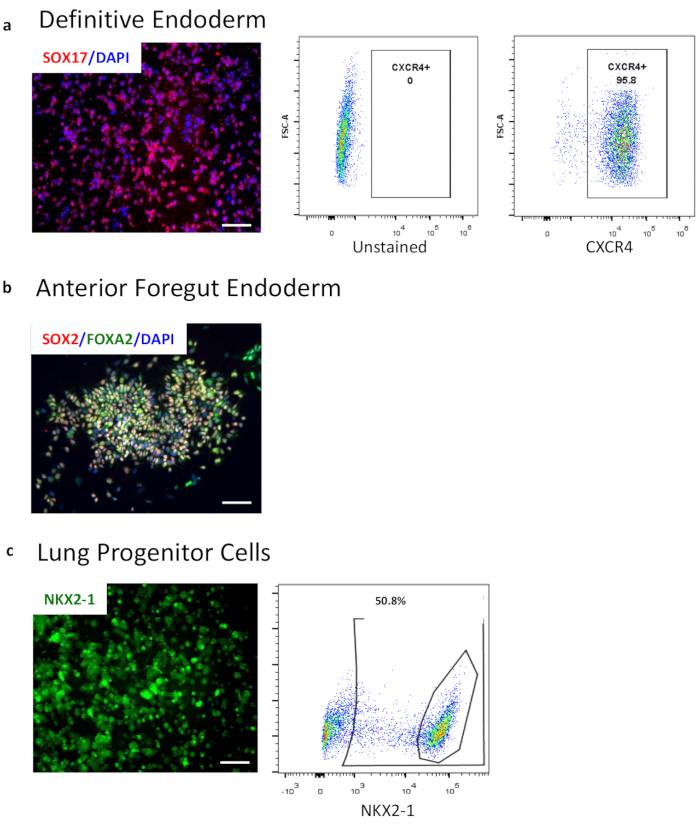
Figure 2: Characterization of endoderm differentiation by flow cytometry and immunocytochemistry. (a) Flow cytometry of definitive endoderm marker CXCR4. Left panel shows the gating against the unstained population while the middle panel shows the CXCR4 positive population. The right panel shows immunocytochemistry image of SOX17 (red) overlaid with nuclei (blue). (b) Immunocytochemistry image of AFE markers FOXA2 and SOX2 overlaid with nuclei (blue). (c) Endogenous expression of NKX2-1-GFP in a reporter cell line in 3D LPC. Images taken from live cell culture in brightfield and GFP. Flow cytometry of lung progenitor intracellular marker NKX2-1 after fixation and permeabilization. Scale bar size = 50 µM. Please click here to view a larger version of this figure.
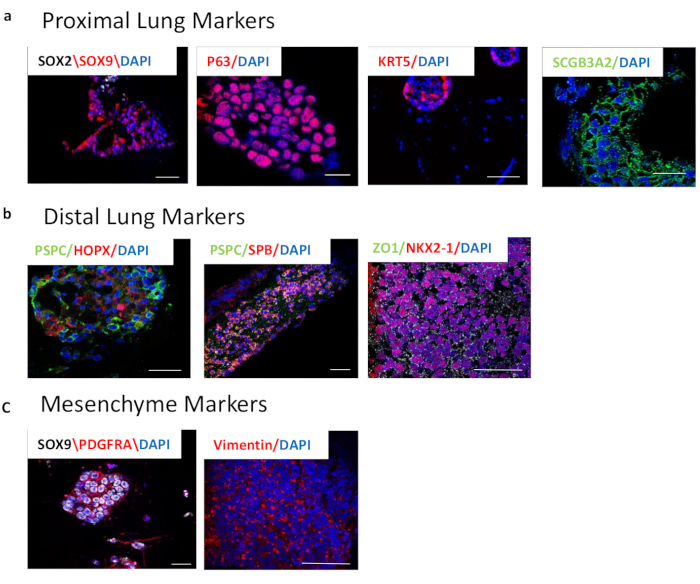
Figure 3: Characterization of 3D whole lung organoids after 3-week differentiation by immunocytochemistry. (a) Proximal lung markers. Left panel shows SOX2 (white) and SOX9 (red) overlaid by nuclei (blue). These markers are important in branching morphogenesis and represent the proximal and distal epithelial populations. Middle panels show P63 (red) and KRT5 (red), both markers of basal cells. The right panel shows SCGB3A2, a marker of club cells. (b) Distal lung markers. Left panel depicts pro-SPC (PSPC), (green) and HOPX (red), markers of alveolar type II ad I cells, respectively, overlaid with nuclei (blue). Middle panel shows pro-SPC (PSPC) (green) and SPB (red), markers of alveolar type II cells, overlaid with nuclei (blue). The right panel shows NKX2-1 (red) and ZO1 (green) overlaid with nuclei (blue). (c) Markers of lung mesenchyme. Left panel shows PDGFRA (red) and SOX9 (white), representing distal mesenchyme. Right panel shows Vimentin (red), which is dispersed throughout the lung. Scale bar size = 50 µM. Please click here to view a larger version of this figure.
| REAGENTS AND SOLUTIONS – For company names please see the Table of Materials List |
| 3D organoid induction medium (day 17-22) |
| Serum-free basal medium (see recipe) supplemented with: |
| FGF7 (10 ng/mL) |
| FGF10 (10 ng/mL) |
| CHIR99021 (3 μM) |
| EGF (10 ng/mL) |
| 3D organoid branching medium (day 23-28) |
| Serum-free basal medium (see recipe) supplemented with: |
| FGF7 (10 ng/mL) |
| FGF10 (10 ng/mL) |
| CHIR99021 (3 μM) |
| All-trans retinoic acid (0.1 μM) |
| EGF (10 ng/mL) |
| VEGF/PIGF (10 ng/mL) |
| 3D organoid maturation medium (day 29-34) |
| Serum-free basal medium (see recipe) supplemented with: |
| Dexamethasone (50 nM) |
| Br-cAMP (100 μM) |
| IBMX (100 μM) |
| AFE induction medium (day 4-6) |
| Serum-free basal medium (see recipe) supplemented with: |
| SB431542 (10 μM) |
| Dorsomorphin (2 μM) |
| DE induction medium (day 1-3) |
| 48.5 mL RPMI1640 + Glutamax |
| 1 mL B27 without retinoic acid |
| 500 μl HEPES (1%) |
| 500 μl pen/strep |
| Human activin A (100 ng/mL) |
| CHIR99021 (5 μM) – only in the first 24 hours |
| LPC induction medium (day 7-16) |
| Serum-free basal medium (see recipe) supplemented with: |
| BMP4 (10 ng/mL) |
| All-trans retinoic acid (RA) (0.1 μM) |
| CHIR99021 (3 μM) |
| Quenching medium |
| 49 mL DMEM/F12 |
| 1 mL FBS |
| Serum-free basal medium (SFBM) |
| 375 mL Iscove’s Modified Dulbecco’s Medium (IMDM) + Glutamax |
| 125 mL Ham’s F12 |
| 5 mL B27 without retinoic acid |
| 2.5 mL N2 |
| 500 μl ascorbic acid, 50 mg/mL |
| 13 μl monothioglycerol/1ml of IMDM” use 300ul of 0.4mM monothioglycerol per 100ml of serum free media |
| 3.75 mL bovine serum albumin (BSA) Fraction V, 7.5% solution |
| 500 μl pen/strep |
| Stem cell passaging medium (day 0) |
| 500 mL DMEM/F12 |
| 129 mL Knockout serum replacement (KSR) |
| 6.5 mL Glutamax |
| 6.5 mL NEAA |
| 1.3 mL 2-mercaptoethanol |
| 6.5 mL pen/strep |
Table 1: Table of media.
| Problem | Solution |
| DE differentiation not efficient | 24 hours after plating in stem cell medium, cells should be 50-70% confluent |
| GSK3β inhibitor/Wnt activator CHIR99021 should be removed within 20-24 hours of day 1 DE induction | |
| DE differentiation should not exceed a total of 72 hours | |
| AFE differentiation not efficient | Ensure that DE differentiation was successful and the cells express > 80% CXCR4 |
| Ensure fresh growth factors/small molecules are being added to the media daily | |
| LPC differentiation not efficient | Ensure the AFE differentiation was successful and the cells express > 80% FOXA2/SOX2 |
| Ensure the AFE cells are passaged as aggregates of 4-10 cells and not single celled | |
| 3D lung organoids not growing or differentiating | Ensure the LPC differentation was successful and the cells express > 30% NKX2-1 |
| Ensure the LPCs were passaged as aggregates of 4-10 cells and not single celled | |
| Ensure there is no residual matrigel from the LPCs during passaging | |
| Add ROCK Inhibitor Y-27632 with each media change | |
| Ensure the media is changed on time and fresh growth factors/small molecules are added | |
| Ensure concentration of growth factors/small molecules is correct |
Table 2: Troubleshooting.
