Canton S (Cs) was used as a wild-type strain. Flies were maintained on a 12 h light: 12 h dark schedule, and sleep deprived for 12 hours overnight. Inspection of the sleep profiles of Cs flies on the baseline day (bs), sleep deprivation day (sd), and two recovery days (rec1 and rec2) (Figure 2A) suggests that flies were effectively sleep deprived in the SNAP, and recovered sleep during the day consistent with observed reports in the literature4,5. The effectiveness of the SNAP in keeping flies awake is also seen in the high activity (300-350 counts/h) exhibited by flies during sleep deprivation (Figure 2B). Indeed, monitoring the activity counts of flies during sleep deprivation can be a useful barometer of the effectiveness of the deprivation protocol and/or an indirect measure of sleep drive. When the sleep deprivation is ineffective, flies are not as active during the period of deprivation. Flies that are under high sleep drive quickly fall asleep after each stimulus and do not traverse the tube as much35. Both the angle of tilt of the apparatus, and the speed of the drop are critical to ensuring that flies are effectively kept awake without harming them. Each lab can optimize the angle and velocity by adjusting the spring (Figure 1B) and/or the size and shape of the cam (Figure 1C and Figure 1D, right).
To quantitatively estimate the effectiveness of sleep deprivation and of recovery, sleep lost during deprivation and then regained in the recovery days was calculated for each individual fly (Figure 2C). Importantly, there was no significant change in baseline sleep between the deprivation day and the baseline day (see 0-12 h in Figure 2C) indicating that sleep is stable in these flies. A large difference in sleep in this 12 hour period (e.g., ± 100 min) would suggest that sleep was not stable. The SNAP effectively deprived flies of >98% of their night-time sleep. Flies recovered ~20% of their sleep in the first 12 h and did not recover additional sleep during the night, as previously reported. However, flies began to recover sleep the following day such that they recovered ~36% of their sleep over 48 h of recovery (Figure 2D). 30 – 40% recovered sleep over 48 h is fairly typical for wild-type flies sleep deprived using the SNAP.
Sleep homeostasis is characterized both by increased sleep duration and by increased sleep depth during the recovery period following deprivation. Daytime sleep consolidation is commonly used as a readout of sleep depth. Sleep consolidation can be assessed as the average sleep bout duration over the entire day (Figure 2E). However, as sleep pressure is dissipated during recovery, the average sleep bout duration will be reduced as the day progresses. Thus, it is frequently helpful to also examine changes in the maximum sleep bout duration which can provide a more sensitive metric (Figure 2F).
| Method of Sleep Deprivation | Total # of papers | % papers / technique | Avg recovery evaluated |
| SNAP | 52 | 37.14% | 33 ± 3 |
| Vortexer/Random Shaking | 49 | 35.00% | 18 ± 3 |
| Hand-Deprivation | 9 | 6.43% | 36 ± 11 |
| Thermogenetic SD | 15 | 10.71% | 36 ± 12 |
| Unspecified | 15 | 10.71% | 29 ± 10 |
Table 1: Survey of different methods of sleep deprivation used in the literature. Only 116 /254 papers used sleep deprivation. The number of papers using each method = "Total # of papers". The fraction of papers using each method = "% papers / technique". The mean length of recovery evaluated for each method = "Avg recovery evaluated". SD – Sleep deprivation. SNAP – Sleep Nullifying Apparatus
| Length of SD | Total studies |
| < 6 h | 12 |
| 6 h | 23 |
| >6 h & < 12 h | 17 |
| 12 h | 69 |
| >12 h & <24 h | 7 |
| 24 h | 19 |
| > 24 h | 9 |
| Chronic SD | 4 |
| Any SD | 160 |
Table 2. Length of sleep deprivation performed in different studies. SD – Sleep deprivation

Figure 1. The Sleep Nullifying APparatus (SNAP). A) Front view of the apparatus. The SNAP can accommodate 8 activity monitors in two rows; holder pins restrain the monitors in place. The legs can be adjusted to help position the apparatus at the correct orientation. B) Closeup view of the motor and spring that rock the apparatus back and forth. The motor turns a cam that tilts the apparatus back to the "up" position and compresses the spring. Release of the spring from compression snaps the apparatus back to the "down" position. C) Left – Side view of the apparatus in the "down" position. Holder pins restrain monitors; a monitor cord slot ensures that monitor cords are held in place. Pads help cushion the impact of the apparatus snapping to the 'down' position. Right – Close view up of the cam. D) Left Side view of the apparatus in the "up" position. Right – The counter-clockwise rotation of the cam tilts the apparatus into the 'up' position. Please click here to view a larger version of this figure.
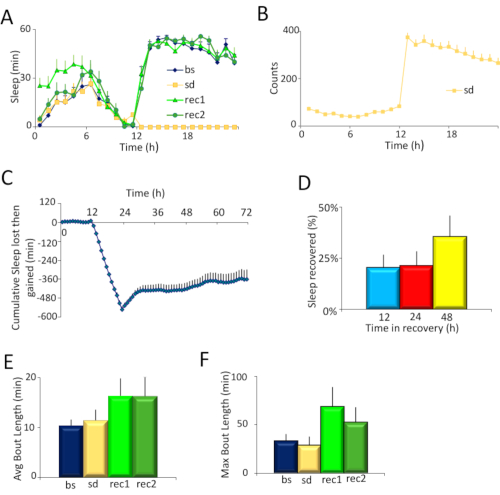
Figure 2. Experimental results. A) Sleep plots of Cs flies for the four days of the experiment: the baseline day (bs), sleep deprivation day (sd), and two days of recovery (rec1 and rec2). B) Average locomotor activity counts of flies on the day of sleep deprivation. Flies were sleep deprived from hours 12-24. C) Time course of sleep deprivation and recovery. Cs flies were sleep deprived from hours 12 – 24, and allowed to recover from hour 24 – 72. The SNAP effectively deprived flies of >98% sleep, which was partially recovered over 48 h (n = 12 flies, Repeated Measures ANOVA for time, F [70,1470]=12.97, p < 10-15). D) Percentage of sleep recovered over 48 h. Flies recovered ~20% of their sleep over 12 h, and ~36% of their sleep over 48 h. E) Sleep consolidation for each day of the experiment as measured by average sleep bout duration during the day. Sleep is more consolidated on the first recovery day compared to baseline (p <0.05, t-test). F) Sleep consolidation for each day of the experiment as measured by maximum sleep bout duration during the day. Sleep is more consolidated on the first recovery day compared to baseline (p <0.05, t-test). Please click here to view a larger version of this figure.
