Here, a worm expressing both GCaMP and TagRFP under control of the AVA interneuron-specific promoter is used as an example of calcium imaging in freely behaving C. elegans. GCaMP and TagRFP channel data were obtained as a series of images, some of which are shown in Figure 6 and as a movie (Supplemental Movie 1). The displacement of the Petri plate induced by our nonlocalized vibration system (Figure 7) was also quantified. The displacement can be controlled by setting the amplitude value in the software for vibration control (Figure 3B) and the Volume adjuster in the amplifier (Figure 4), whereas the frequency is regulated by setting the frequency value in the software (Figure 4). In the successful experiments, a transient calcium response of AVA neurons was observed upon stimulation with vibrations having a frequency of 630 Hz and a displacement of approximately 4.5 µm for 1 s, indicating that the AVA neuron was activated during a worm's backward response to the nonlocalized stimulation (Figure 6). Baselines of both GCaMP and TagRFP signals also showed that almost no photobleaching occurred during recording.
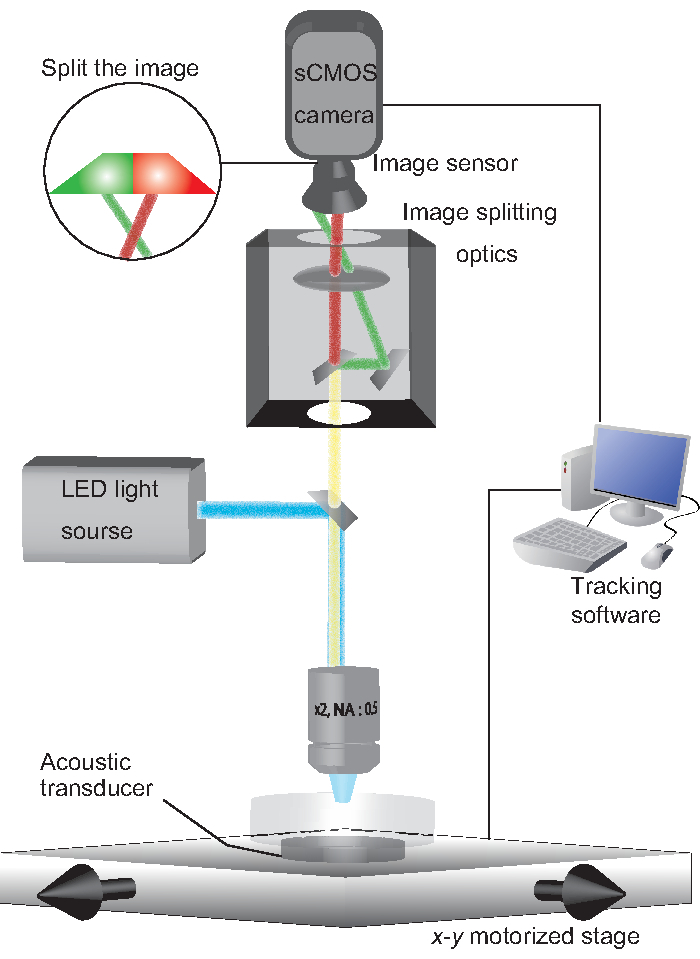
Figure 1: A schematic system configuration. An NGM plate on which a worm freely moves is held on an acoustic transducer. Excitation light (488 nm and 560 nm) is emitted from an LED light source. GCaMP and TagRFP fluorescent emissions are split by image-splitting optics, such that each fluorescent emission is projected onto half of an sCMOS camera. Tracking software controls the x-y motorized stage to track the fluorescent spot of the worm. Please click here to view a larger version of this figure.
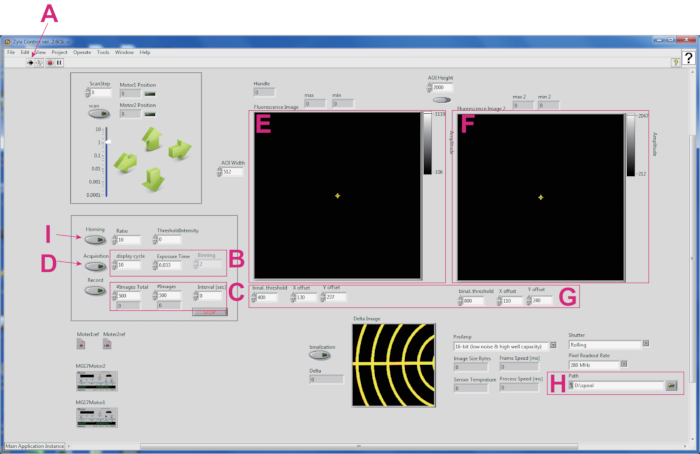
Figure 2: Screenshot of the software for tracking a fluorescent spot of a worm. (A) The Run button for activating the software. (B) Boxes for setting image display cycle, exposure time (s), and binning in each channel. (C) Boxes for providing information on the image acquisition cycle. (D) The Acquisition button to initiate live streaming. (E) GCaMP image. (F) TagRFP image. (G) A panel for calibration coordinates between GCaMP and TagRFP images. (H) Box for setting file path. (I) The Homing button to start worm tracking. Please click here to view a larger version of this figure.
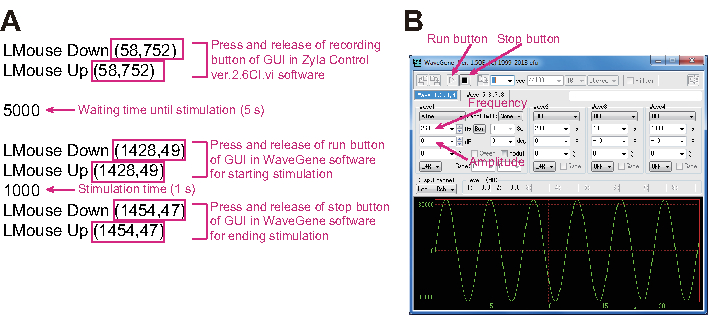
Figure 3: Photo of the mouse macro system and software for controlling vibration. (A) The magenta square indicates the coordinates for the mouse cursor. The meaning of each script line is explained in magenta letters. (B) The GUI (graphical user interface) of software for controlling vibration. Vibration frequency and amplitude values are controlled using this GUI. Please click here to view a larger version of this figure.

Figure 4: Photo of the NGM plate held on the acoustic transducer using transparent adhesive tape. Only a single worm is moving on the plate. Please click here to view a larger version of this figure.
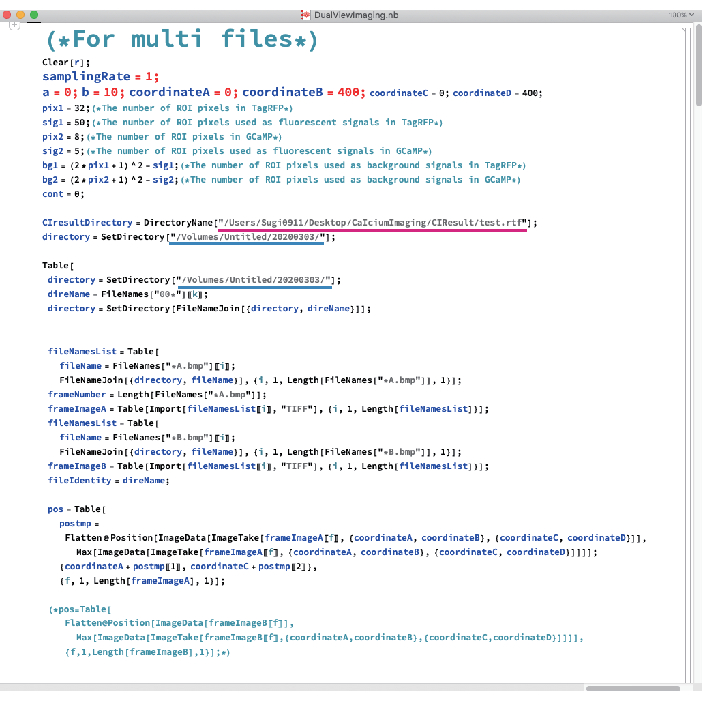
Figure 5: Photo of the data analysis software. Magenta and blue underlines indicate the paths to the CIResult folder and the folder including BMP files, respectively. Please click here to view a larger version of this figure.
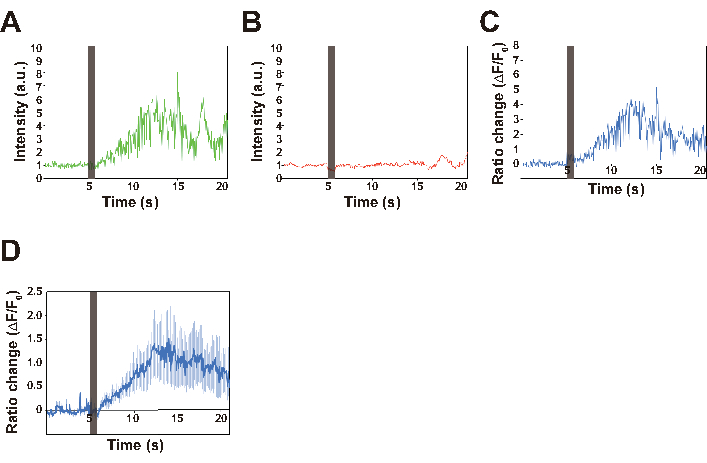
Figure 6: Traces of the intensities of GCaMP (A), and TagRFP (B) fluorescence, and the ratio change (C,D). (A–C) The representative traces for GCaMP signal, TagRFP signal and ratio changes of a single worm. The ratio change is calculated using the DualViewImaging.nb file, as follows. The coordinate in which the intensity of TagRFP fluorescence is maximum after background subtraction is extracted for each image. This TagRFP signal serves to compensate for focus changes caused by the animal's motion. The maximum fluorescence intensity of GCaMP within the extracted coordinate ±10 pixels is calculated as the GCaMP signal intensity for each image. For ratiometry, ((IGCaMP − IGCaMP_BG) − (ITagRFP − ITagRFP_BG)) / (ITagRFP − ITagRFP_BG) is calculated, in which IGCaMP and ITagRFP are the GCaMP and TagRFP signals from the AVA neuron, respectively, and IGCaMP_BG and ITagRFP_BG are the background signals, respectively. This calculation process was applied to all images to quantify the ratio change in a reversal event. The vertical line at 5 s in all traces indicates the stimulation period. (D) The average ratio changes for 15 worms. Error bar indicates SEM. Please click here to view a larger version of this figure.
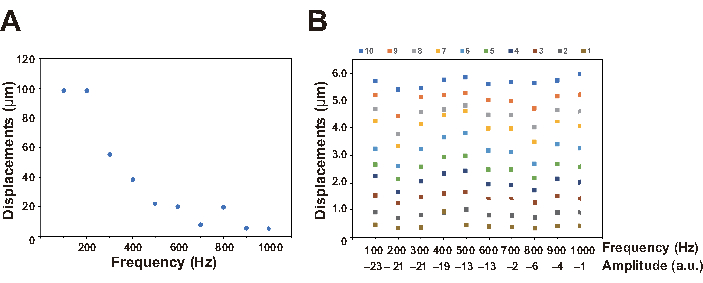
Figure 7: Relationship between parameters. (A) Replacements were measured every 100 Hz using a laser doppler vibrometer, in which the amplitude value in the software and the VOLUME adjuster value in the amplifier were fixed to 0 and 10, respectively. The resonance frequency was observed between 100-200 Hz, which was similar to the value described in the specification of the acoustic transducer. (B) Frequencies and displacements were controlled by the software (range; 100 to 1000 Hz) and the amplifier (range: 1-10), respectively and measured using a laser doppler vibrometer. The amplitude value selected by the software is indicated for each frequency. Please click here to view a larger version of this figure.
Supplementary Movie 1: A representative movie for calcium transients in an AVA interneuron of a freely behaving worm in response to a 630 Hz nonlocalized vibration. Stimulation was evoked 5 s after starting the recording. GFP was also expressed in the intestine of the worm as a co-injection marker for GCaMP. This GFP fluorescence does not affect the signal intensity of GCaMP. The scale bar is also indicated in the first image. The direction of the worm is indicated at the top right after the first image. The Windows media player is suitable for playing the AVI file, but it can also be played using some free downloaded media players, such VLC media player in the Mac. Please click here to download this File.
